

서언
재료 및 방법
실험재료
추출물 제조
세포 생육 억제활성 측정
SDS-PAGE 및 Western blot 분석
Reverse transcriptase-polymerase chain reaction (RT-PCR)
통계분석
결과 및 고찰
STB와 STL의 SW480과 A549세포에 대한 세포생육억제 활성
STB와 STL의 SW480과 A5490세포에서 β-catenin 단백질 및 유전자 발현 조절 활성
STB와 STL의 SW480과 A5490세포에서 β-catenin 단백질 분해 유도 활성
STB와 STL의 SW480과 A5490세포에서 β-catenin 단백질 분해 유도에 있어 GSK3β와 ROS의 영향
적요
서언
Wnt/β-catenin 신호전달경로라고도 하는 Wnt 경로의 주요 단백질인 β-catenin은 다양한 암의 진행과 전이에 관여하는 것으로 알려져 있다(Kypta and Waxman, 2012; Watson et al., 2013). Wnt/β-catenin 신호전달은 Wnt ligands가 Frizzled/low-density lipoprotein receptor related protein receptor 복합체에 결합함으로써 진행되며, GSK3β-Axin-APC destruction 복합체를 불활성화 시켜, β-catenin/T-cell factor 경로를 통해 신호전달을 유도한다(MacDonald et al., 2009). 이러한 Wnt/β-catenin 신호전달은 세포주기, 줄기 세포의 자체 재생 및 전이의 조절을 통해 종양 발생 및 진행에 중요한 역할을 한다(MacDonald et al., 2009; Reya and Clevers, 2005). 핵 내 β-catenin의 비정상적인 축적은 Wnt/β-catenin 신호전달 활성화의 특징으로 핵 내 축적된 β-catenin은 T-cell factor (TCF)/lymphoid enhancer factor (LEF)에 결합하여 MYC과 cyclin D1과 같은 발암유전자의 발현을 유도한다(Chitalia et al., 2013; Xue et al., 2015). 그러므로, Wnt/β-catenin 신호전달경로의 핵심단백질인 β-catenin의 발현을 억제할 수 있는 항암제 개발이 요구되고 있다(Kim and Kim, 2016; Yu et al., 2018).
갈매나무과(Rhamnaceae) 식물인 상동나무(Sageretia thea)는 약용식물로서 한국과 중국에서 간염과 발열을 위한 치료제로 사용되어 왔다(Chung et al., 2009; Hyun et al., 2015). 최근 상동나무의 기능성 연구에서 상동나무 열매는 항산화, 항당뇨 및 미백활성이 있다고 보고되었다(Hyun et al., 2015; Ko et al., 2018). 상동나무 잎은 항산화 활성을 통한 저밀도지단백질의 산화 억제와 항암활성을 나타낸다고 보고되고 있으며(Hyun et al., 2015; Kim et al., 2019; Park et al., 2002), 상동나무 가지는 대장암세포와 유방암세포의 세포사멸을 유도한다고 보고되고 있다(Kim et al., 2019; Ko et al., 2016). 상동나무의 항암활성 관련 작용기전 연구에서 가지와 잎 추출물은 GSK3β 의존성 cyclin D1 인산화를 통해 cyclin D1의 분해 및 Nrf2 의존성 HO-1 발현 유도를 통해 대장암세포의 세포생육억제와 세포사멸을 유도하다고 보고되었다(Kim et al., 2019). 상동나무의 β-catenin 단백질 발현 조절활성에 대한 연구는 미비하여 본 연구에서 상동나무 가지와 잎 추출물의 대장암세포와 폐암세포에서 β-catenin 단백질 발현 조절활성을 조사하였다.
재료 및 방법
실험재료
대장암 및 폐암세포의 배양을 위한 배지인 DMEM/F-12는 Lonza (Morristown, NJ, 에서 구매하였다. 단백질분해효소 억제제인 MG132, GSK3β 억제제인 LiCl, 활성산소제거제인 N-acetyl Cysteine (NAC) 및 3-(4,5-dimethylthizaol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)는 Sigma-Aldrich (St. Louis, MO, USA)사에서 구매하였다. 그리고 western blot 분석을 위한 항체인 β-catenin과 actin은 Cell Signaling Technology (Danvers, MA, USA)에서 구매되었다.
추출물 제조
본 실험에 사용된 시료는 갈매나무과(Rhamnaceae)에 속하는 상동나무(Sageretia thea) 잎과 가지는 국립산림과학원 산림약용자원연구소로부터 제공받았다. 상동나무는 국립산림과학원 산림약용자원연구소의 손호준박사의 식물학적 동정을 거쳤으며, 상동나무의 확증표본(표본번호: FM.J.S. 20180510-01)는 국립산림과학원 산림약용자원연구소에 보관하고 있다. 음건·세절한 상동나무 잎과 가지 40 g을 800 mL의 70% 에탄올을 가하여 상온에서 7일 동안 침지하여 추출하고, 원심분리하여 70% 에탄올 추출물을 회수하였다. 회수된 에탄올 추출물은 40℃ 이하의 중탕에서 감압 환류 냉각장치(N-1110S, EYELA, Tokyo, Japan)로 감압농축 후 동결건조 되었다. 동결건조 된 추출물은 -80℃의 냉동고에 보관하면서 분석을 위한 시료로 사용되었다. 상동나무 가지(STB)와 잎(STL) 추출물은 dimethyl sulfoxide (DMSO)에 용해하여 실험에 사용되었고, 대조군은 DMSO를 0.1%를 초과하지 않는 범위에서 사용하였다.
세포 생육 억제활성 측정
본 연구에서 사용된 대장암세포 SW480과 폐암세포 A549는 American Type Culture Collection (Manassas, VA, USA)에서 구매되었고, 각각의 세포는 10% FBS, 100 U/mL penicillin과 100 ㎍/mL streptomycin이 포함된 DMEM/F-12 배지로 37℃, 5% CO2 하에서 배양되었다. SW480과 A549세포의 생육억제는 MTT assay로 측정하였다. SW480과 A549세포를 96-well plate에 well 당 1 × 104 cells로 24 시간 배양한 후, STB와 STL을 농도별 처리하고 24시간 동안 배양시켰다. 24시간 후 각각의 well에 MTT 용액(1 ㎎/mL)을 50 μL씩 첨가하고 2시간 반응시킨 후, 상등액을 제거하고 DMSO를 100 μL씩 각각의 well에 첨가하여 녹인 후 UV/Visible spectrophotometer (Xma-3000PC, Human Corporation Co., Seoul, Korea)를 이용하여 540 ㎚에서 흡광도를 측정하였다.
SDS-PAGE 및 Western blot 분석
세포로부터 단백질을 추출하기 위해, 처리 후 세포를 4℃에서 유지된 1 × phosphate-buffered saline (PBS)로 2회 세척한 후, protease inhibitor cocktail (Sigma-Aldrich Co.)과 phosphatase inhibitor cocktail (Sigma-Aldrich Co.)이 포함된 NP-40 lysis buffer (GenDEPOT, Katy, TX, USA)를 4℃에서 1시간 처리하여 단백질을 추출하였다. 추출된 단백질은 Bicinchoninic acid protein assay (Pierce Biotechnology Inc., Waltham, MA, USA)로 정량분석 후, 동일량의 단백질을 8% SDS-acrylamide gel로 전기영동하고 PVDF membrane (Bio-Rad, Hercules, CA, USA)에 이동시킨 후 5% non-fat dry milk로 상온에서 1 시간 동안 blocking 하였다. 1시간 후, 1 차 항체를 5% non-fat dry milk에 용해시켜 4℃에서 16시간 동안 반응시킨 후 membrane을 0.05% tween-20이 포함된 tris-buffered saline (TBS-T)로 5분간 3회 세척하였다. 그 후 2 차 항체는 5% non-fat dry milk에 용해시켜 membrane에 상온에서 1시간 처리하였고, TBS-T로 5분간 3회 세척 후 membrane은 ECL western blotting substrate (Amersham Biosciences Co., Little Chalfont, England)를 이용하여 단백질을 확인하였다.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
세포에서 RNA를 추출하기 위해서 세포를 4℃에서 유지된 1 × PBS로 2회 세척한 후, RNeasy Mini kit (QIAGEN GmbH., Hilden, Germany)를 이용하여 total RNA를 추출하였다. 1 ㎍의 total RNA를 Verso cDNA synthesis kit (Thermo Fisher Scientific Inc., Waltham, MA, USA)를 이용하여 cDNA를 합성하였다. PCR은 PCR master mix kit (Promega Co., Madison, WI, USA)를 이용하여 수행되었고, 사용된 primer는 Table 1과 같다.
Table 1. Sequence of oligonucleotide primers used for RT-PCR
| Gene Name | Sequence |
| β-catenin |
Forward: 5'-cccactaatgtccagcgttt-3' Reverse: 5'-aatccactggtgaaccaagc-3' |
| GAPDH |
Forward: 5'-acccagaagactgtggatgg-3' Reverse: 5'-ttctagacggcaggtcaggt-3' |
통계분석
모든 결과는 3 회 반복 측정 후 평균 ± 표준편차로 나타내었고, 처리간 유의성은 Student’s t-test로 검증하여 p-value 값이 0.05 미만일 때 통계적으로 유의하다고 판정하였다(Microsoft Exel 2010, Microsoft, Redmond, WA, USA).
결과 및 고찰
STB와 STL의 SW480과 A549세포에 대한 세포생육억제 활성
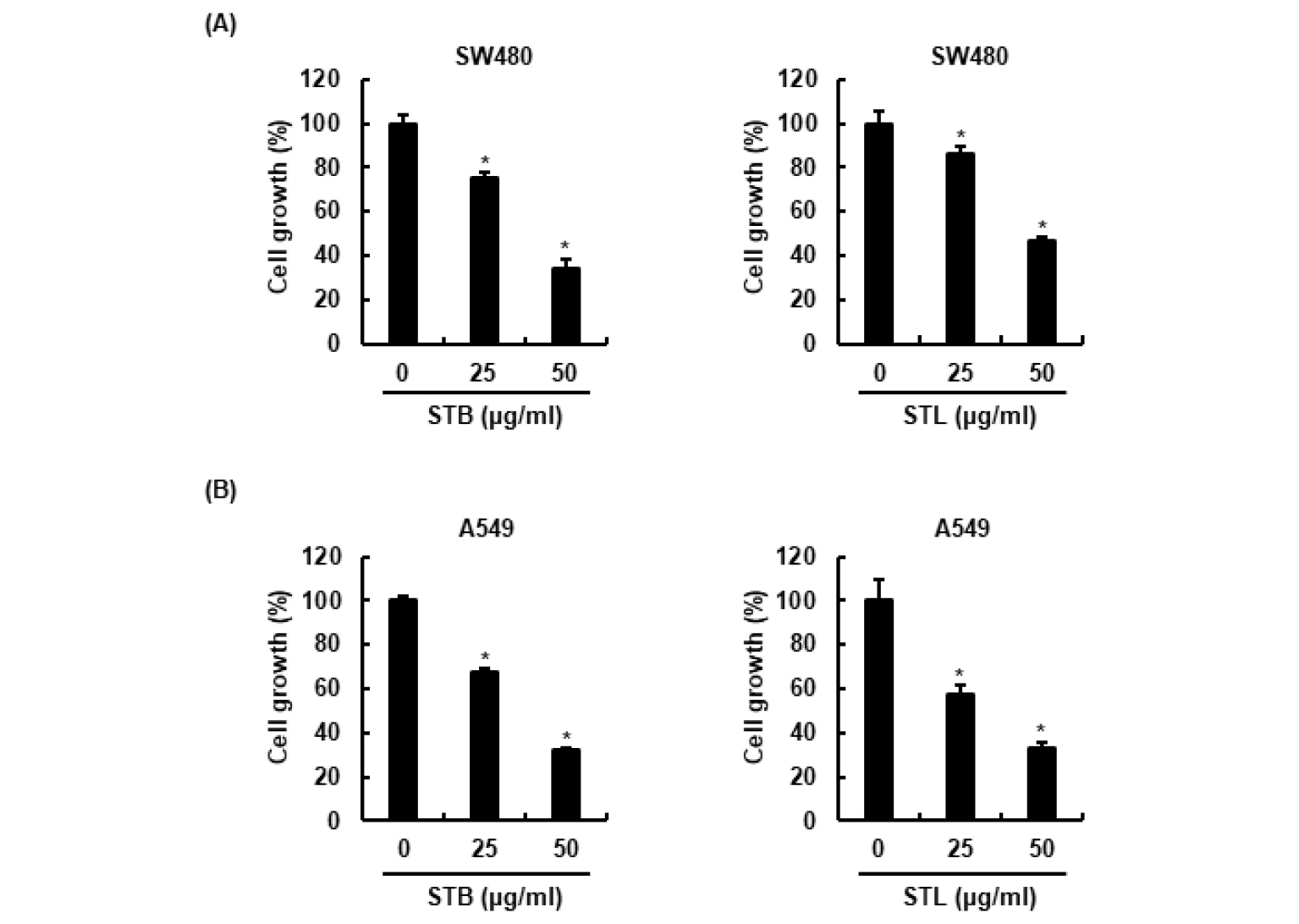
STB와 STL의 대장암과 폐암세포에 대한 세포생육억제 활성을 평가하기 위해, 대장암세포인 SW480과 폐암세포인 A549에 STB와 STL을 농도별로 24시간 처리 후, MTT assay를 이용하여 STB와 STL의 세포생육억제활성을 평가하였다. 그 결과, STB와 STL은 SW480과 A549세포의 세포생육을 농도 의존적으로 억제하는 것으로 나타났다(Fig. 1).
STB와 STL의 SW480과 A5490세포에서 β-catenin 단백질 및 유전자 발현 조절 활성
β-catenin은 생리적 항상성에 중요한 역할을 한다고 알려져 있지만, 비정상적인 β-catenin의 발현은 암을 비롯한 다양한 질병을 초래한다고 보고되고 있다(Shang et al., 2017). 비정상적인 β-catenin의 발현은 cyclin D1과 c-Myc과 같은 발암단백질의 발현을 유도하여 종양형성과 암세포의 증식을 초래한다(Damsky et al., 2011; Gekas et al., 2016; Khramtsov et al., 2010; Kobayashi et al., 2000; Tao et al., 2014; Valkenburg et al., 2011). 실제적으로, 비정상적인 β-catenin의 발현이 대장암, 간암, 췌장암, 폐암 및 난소암의 발암 및 종양형성을 초래한다고 알려져 있다(Shang et al., 2017).
STB와 STL의 대장암세포와 폐암세포의 β-catenin 발현 조절활성을 단백질 및 mRNA 수준에서 평가한 결과, STB와 STL은 A549와 SW480세포에서 β-catenin의 단백질 및 mRNA 수준을 감소시켰으나 STB와 STL에 의해서 유도되는 β-catenin 단백질 감소가 mRNA 감소에 비해 급격하게 나타났다(Fig. 2A and 2B). 본 결과를 미루어 볼 때, STB와 STL은 β-catenin mRNA 발현 억제를 통해 β-catenin 단백질을 감소시키고, 또한 β-catenin 단백질의 분해를 유도하는 것으로 판단된다.
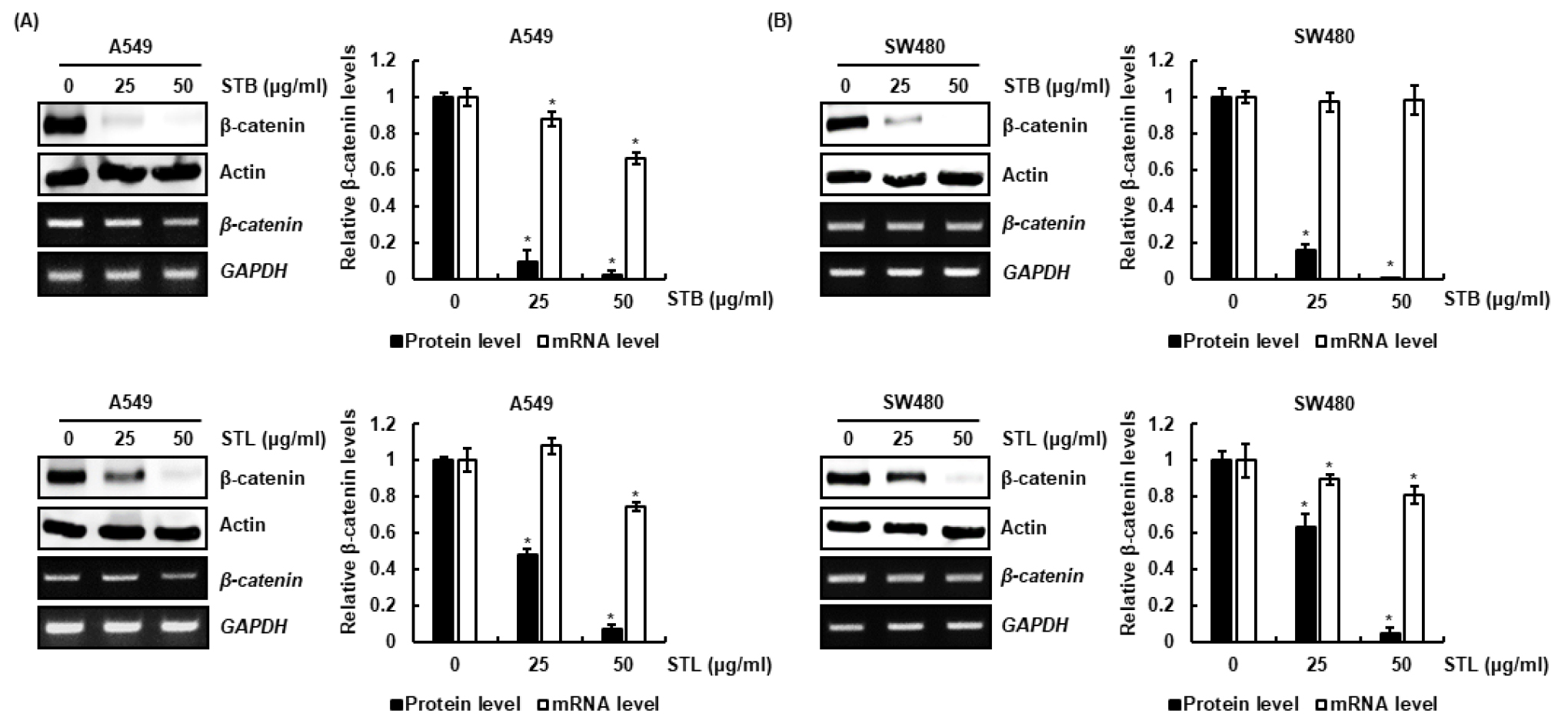
Fig. 2.
Effect of STB and STB on β-catenin expression in SW480 and A549 cells. (A and B) A549 and SW480 cells were treated with STB and STL at the indicated concentrations for 24 h. Western blot and RT-PCR analysis was performed against β-catenin. Actin and GAPDH were used as internal controls for Western blot and RT-PCR, respectively. *p < 0.05 compared to cell without STB and STL.
STB와 STL의 SW480과 A5490세포에서 β-catenin 단백질 분해 유도 활성
비 정상적인 Wnt 신호전달은 β-catenin 단백질 분해 유도 복합체인 GSK3β-Axin-APC destruction 복합체를 불활성화시켜 β-catenin 단백질의 안정화를 유도하는데, 이것은 종양형성에 중요한 역할을 한다고 알려져 있다(Johnson et al., 2005; Morin et al., 1997). GSK3β-Axin-APC destruction 복합체가 불 활성화됨으로써 β-catenin 단백질은 세포질에 축적되고, 결국에는 핵으로 전위되어 TCF/LEF와 결합하여 암세포의 증식관련 유전자의 전사를 유도한다(White et al., 2012). 따라서, β-catenin 단백질 분해 유도는 항암제 개발의 주요 표적으로 여겨지고 있다. 실제적으로, Celecoxib, DIF1/3, Genistein, G007-LK, XAV939, JW55 및 Pyrvinium과 같은 항암분자들이 β-catenin 단백질 분해를 유도하는 것으로 보고되었다(Shang et al., 2017).
STB와 STL이 β-catenin 단백질 분해를 유도하는지 평가하기 위해, 단백질 분해효소 억제제인 MG132를 2시간 동안 처리한 후, STB와 STL을 12시간 처리하여 β-catenin 단백질을 확인한 결과, MG132가 처리되지 않은 SW480과 A5490세포에서는 STB와 STL에 의한 β-catenin 단백질 감소가 확인되었으나, MG132가 처리된 세포에서는 β-catenin 단백질 감소가 억제되었다(Fig. 3A and 3B). 본 결과를 미루어 볼 때, STB와 STL은 β-catenin 단백질의 분해를 유도하는 것으로 사료된다.
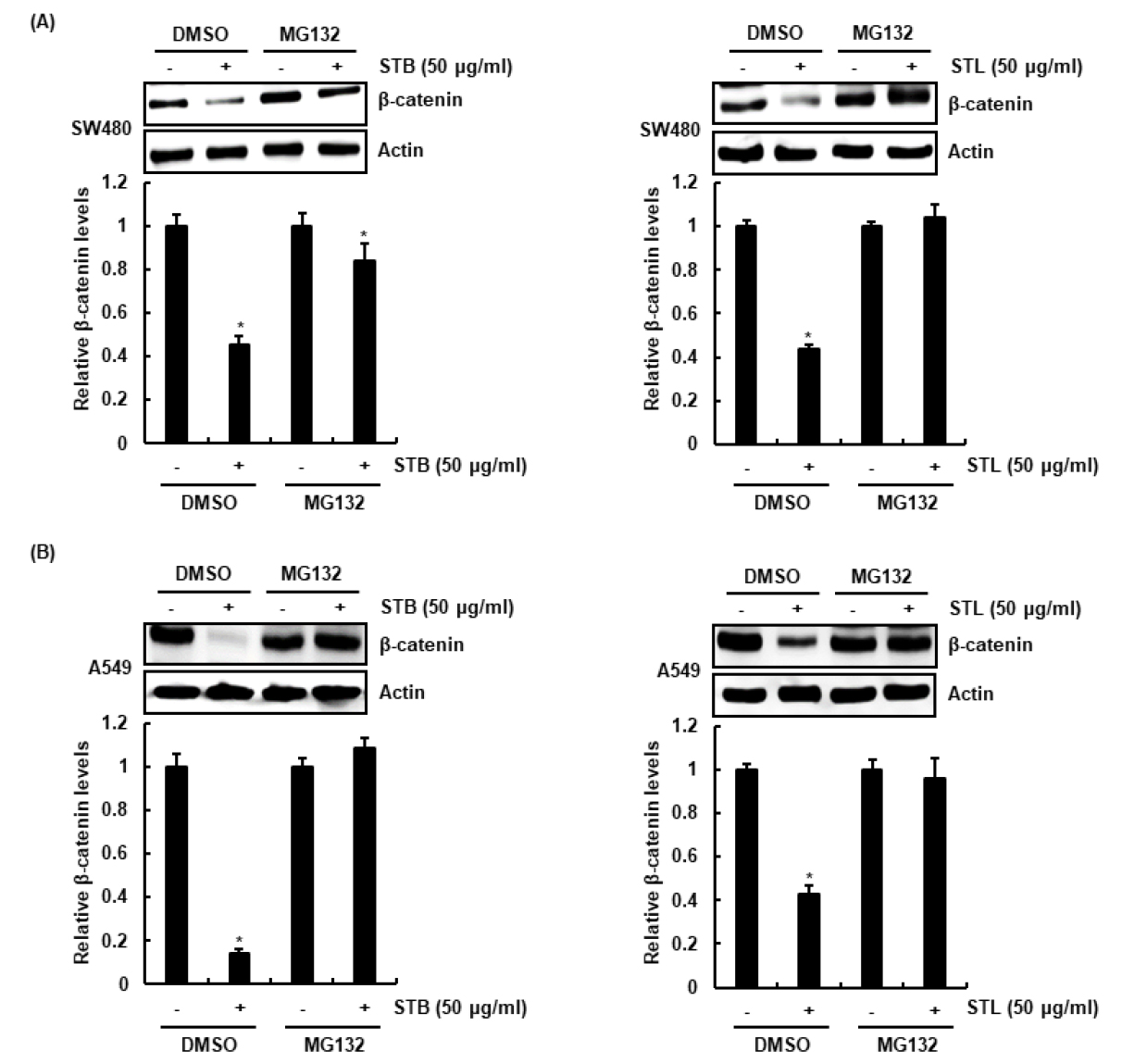
Fig. 3.
Effect of STB and STL on β-catenin proteasomal degradation in SW480 and A549 cells. (A and B) SW480 and A549 cells were pretreat with 20 μM of MG132 for 2 h and then co-treated with STB and STL for 12 h. Western blot analysis was performed against β-catenin. Actin was used as an internal control. *p < 0.05 compared to cell without STB and STL.
STB와 STL의 SW480과 A5490세포에서 β-catenin 단백질 분해 유도에 있어 GSK3β와 ROS의 영향
β-catenin 단백질 분해는 GSK3β에 의한 β-catenin의 인산화와 관련이 있다고 알려져 있다(Li et al., 2012). GSK3β에 β-catenin의 아미노말단의 threonine-41, serine-37과 serine-33이 인산화되면 F-box-containing protein인 β-TrCP ubiquitin E3 ligase에 의해 유비퀴틴화가 되어 proteasome에 의해 β-catenin이 분해된다(Li et al., 2012). 또한, ROS는 caspase-3를 활성화시켜 β-catenin의 분해를 유도한다고 보고되었다(Omori et al., 2011). 그리하여, STB와 STL에 의해 야기되는 β-catenin 분해 유도에 GSK3β와 ROS가 관여하는지 검증하기 위해, GSK3β 억제제인 LiCl과 ROS 제거제인 NAC를 처리하고, STB와 STL을 12시간 처리하여 β-catenin 단백질을 확인하였다. 그 결과, LiCl과 NAC가 처리되지 않은 세포와 처리된 세포에서 모두 STB와 STL에 의한 β-catenin 단백질 감소가 나타났다(Fig. 4A and 4B). 본 결과를 미루어 볼 때, STB와 STL에 의해 유도되는 β-catenin 단백질 분해는 GSK3β와 ROS 비의존성인 것으로 사료된다.
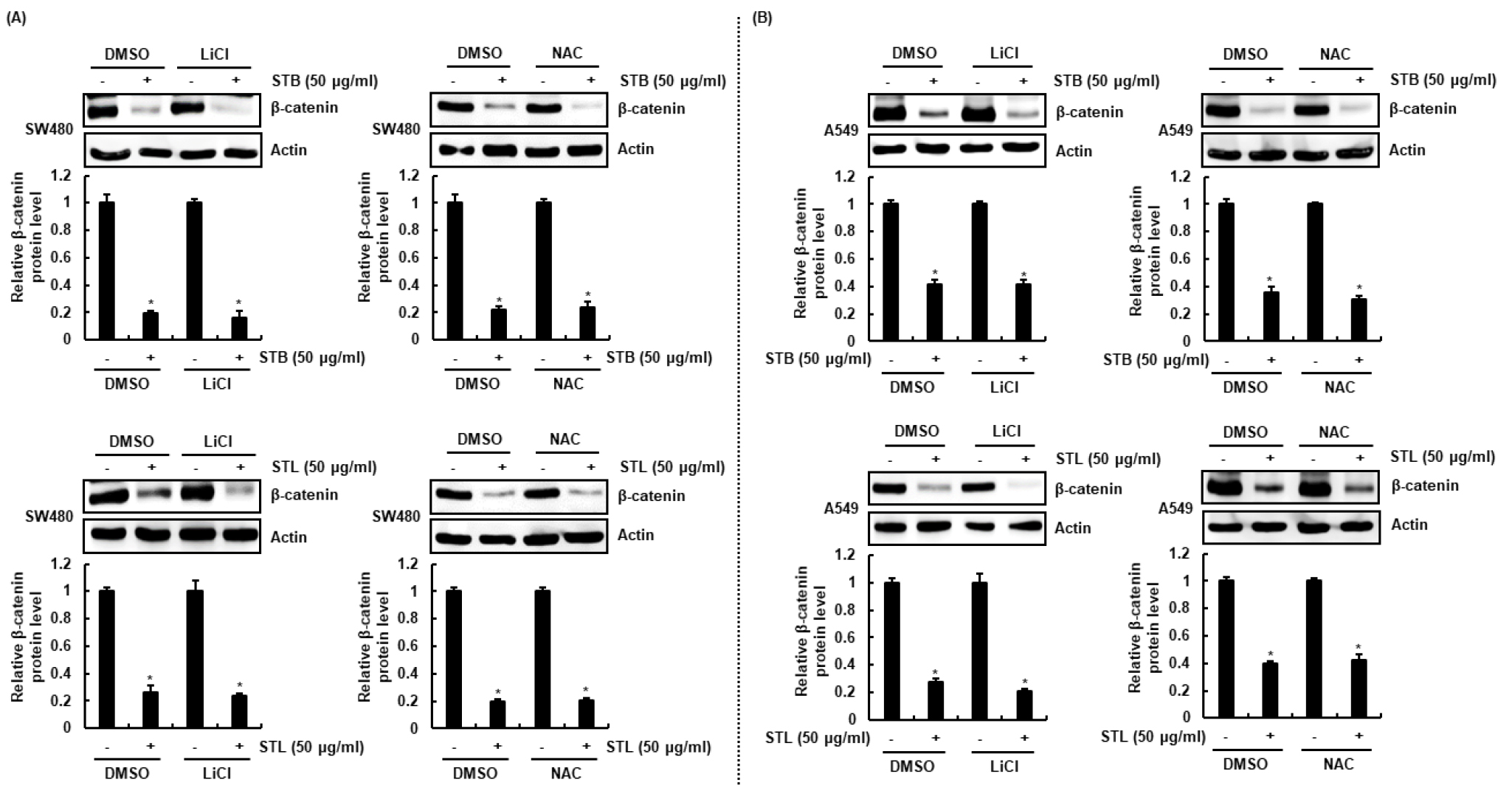
Fig. 4.
Effect of GSK3β or ROS on β-catenin proteasomal degradation induced by STB and STL in SW480 and A549 cells. (A and B) SW480 and A549 cells were pretreated with 20 mM of LiCl or 10 mM of NAC for 2 h and then co-treated with STB and STL for 12 h. After the treatment, Western blot analysis was performed against β-catenin. Actin was used as internal control. *p < 0.05 compared to cell without STB and STL.
