

서언
갈매나무과 대추나무속에 속하는 대추(Ziziphus jujuba)는 중국이 원산지로 중국 및 한국을 포함한 아시아, 유럽, 미국, 캐나다, 오스트레일리아 등 30여개국에서 재배되고 있다(Liu, 2003). 대추는 중국에서 경제적으로 중요한 과일로서 연간 43백만톤이 생산되고 있다(Huang et al., 2016). 한국에서는 2015년 14,199톤의 대추가 생산되었고, 생산액은 980억원이다(KFS, 2016).
핵과류인 대추의 과육에는 당질과 비타민C, 아미노산, 폴리페놀이 풍부하다. 당류는 사탕무 보다 2배, 비타민C는 키위에 비해 약 4배 높다(Li et al., 2007; Liu et al., 2014b). 종자에는 올레산 등 불포화지방산이 포함되어 있다(Park and Kim, 2016). 또한 대추는 항산화작용(Na et al., 1996), 진정작용(Han and Park, 1987) 등 약리효과가 우수하여 건과형태로 한방 재료나 음식 부재료로 다양하게 이용되고 있다.
주로 건과형태의 한방재료로 이용되어온 대추는 2000년대 충북 보은을 중심으로 생대추를 재배하기 시작하며 소비형태가 생과로 바뀌었다. 이후 고소득 작목으로 각광받으며 대추 재배면적이 증가되고 있다(Lee et al., 2017; Oh et al., 2017).
중국에는 건과용, 생과용, 관상용 등 다양한 특성을 가진 700여개의 대추 유전자원이 존재한다(Grygorieva et al., 2014; Huang et al., 2016; Ma et al., 2011). 한국에서는 1980년대에 생과용 품종 ‘무등’, 건과용 품종 ‘금성’, 생과 및 건과 겸용 품종 ‘월출’이 육성되었고, 생과용 대추 생산을 목적으로 2014년에 ‘천황’이 2015년에는 ‘황실’이 육성되었다. 하지만 현재 재배되고 있는 품종은 재래종인 ‘복조’로 국한되어 있으며, 보유하고 있는 대추 유전자원의 다양성이 부족한 실정이다.
다양한 품종 육성을 위해서는 교배육종이 필요하나 대추는 꽃이 1 ㎝ 미만으로 작고 인공수분이 어려우며 단위결과를 하는 특징이 있다(Ackerman, 1961). 또한 한국 보유 대추 유전자원은 종자가 형성되지 않거나 퇴화하는 경우도 많아 충실한 종자를 얻기가 어려워 현재까지 대추 품종 육성은 주로 분리육종이나 아조변이체 선발을 통해 이뤄지고 있는 실정이다. 따라서 국내외 대추 유전자원을 수집하여 특성검정을 통해 형태형질에 대한 데이터베이스를 구축하고 품종 육성소재로 활용 가능한 종자가 형성되며 우수한 특성을 지닌 유전자원의 확보가 중요하다.
대추 주요 재배 국가인 중국에서는 sequenced-related amplified polymorphism (SRAP)와 simple sequence repeat (SSR) 분자표지를 개발하여 중국 대추 유전자원의 유전적 다양성을 분석하였다(Li et al., 2010; Liu et al., 2014a; Wang et al., 2014). 또한 Zhang et al., (2015)은 대추 유전자원의 형태적 특성과 SSR 분자표지를 활용하여 중국의 주요 대추 품종의 분류학적 위치를 확인하였다. Jang et al. (2006)과 Nam (2016)은 한국 대추 유전자원의 가지의 형태, 개화시각, 과형, 엽장 등이 대추 품종을 구분하기 위한 중요 형질이지만 대추 품종 간 형태적 특성이 유사하여 다형성이 크지 않았다고 보고하였다. 또한 대추 품종 육성을 위한 기초 정보인 종자 형성 여부가 확인되지 않아 교배육종의 양친으로 활용 가능한 유전자원 또한 확인되지 않은 실정이다.
최근 충청북도농업기술원 대추연구소에서 보유 중인 대추 유전자원 중 ‘일본’이 다른 유전자원들과 비교하여 형태적 특성이 특이한 것으로 관찰되었다. 따라서 본 연구는 ‘일본’을 포함한 주요 대추 유전자원의 형태 형질을 조사하고 비교하여 대추 유전자원에 대한 기초자료를 제공하고, 실생 획득이 어려운 대추 교배육종의 소재로서 ‘일본’의 가능성을 확인하고자 수행하였다.
재료 및 방법
실험재료
충청북도농업기술원 대추연구소(36°5'77.64"0N, 127°7'48. 32"9E) 포장에 4 × 2 m로 식재된 대추 유전자원 10개를 이용하였다(Table 1). 국내 선발자원인 ‘복조’, ‘무등’, ‘금성’, ‘추석’, ‘보은’과 일본 재래종인 ‘일본’ 및 중국 재래종인 ‘호대추’는 국립원예특작과학원 배연구소에서 수집되었다. ‘보은’은 충청도 재래종이며, ‘티벳왕’, ‘상왕’ 및 ‘국광’은 중국 도입종이다.
Table 1.
List of the 10 jujube accessions used for characterization of ‘Ilbon’
zThe year when the jujube accessions were collected at the origin.
yThe institutions provided jujube accessions to Jujube Research Institute, Chungcheongbuk-do Agricultural Research & Extension Services.
xThe year when the jujube accessions were introduced in Jujube Research Institute, Chungcheongbuk-do Agricultural Research & Extension Services.
wTree age at the time of the experiment.
형태형질 조사
2016년부터 2017년까지 농업과학기술 연구조사분석기준(RDA, 2012)의 과수 공통 생육조사 기준과 대추나무 특성조사요령지침서(NFSV, 2009)에 따라 발아기, 개화기, 엽형질, 과실특성, 핵의 특성을 포함한 형태형질을 1개 유전자원 당 4주의 나무에서 조사하였으며, 임의로 3주를 선발하여 3반복하였다.
발아기는 눈이 1 ㎜ 이상 돌출된 시기를 조사하였고, 개화기는 수관 전체에 꽃이 10% 이상 개화한 시기를 조사하였다. 숙기는 한 수체에서 과실의 과피가 80% 이상 착색된 시기를 조사하였다. 개화성은 7월에 오전 9시부터 오후 6시까지 1시간 간격으로 꽃받침이 갈라지는 시간을 조사하여 오전 개화성과 오후 개화성으로 구분하였다.
엽형질은 7월 중순에 충실한 결과지의 중간에 위치한 잎을 60개씩 채취하여 방안자로 엽장, 엽폭, 엽병장을 측정하고 엽형지수를 계산하였다. 20개의 잎의 평균을 1반복으로 하여 총 3반복으로 조사하였다. 엽형은 타원형, 난원형, 난상피침형으로 구분하였고 기부 형태는 원저, 심장저, 예저로 분류하였다. 거치 형태는 둔거치, 예거치, 복예거치로 분류하였다.
과실이 성숙된 10월 초순에 자원별 무작위로 수집한 과실 30개를 대상으로 과실특성을 조사하였다. 과실종경과 횡경은 버니어 캘리퍼스(CD-15CPX, Mitutoyo, Tokyo, Japan)를 이용하여 측정하였고, 과실종경과 횡경의 비율로 과형지수를 계산하였다. 과중은 전자저울(XT 4200C, Precisa, Dietikon, Switzerland)을 이용하여 측정하였다. 과실 당도는 과즙 1 mL을 추출하여 pocket refractometer (PAL-1, ATAGO, Tokyo, Japan)로 측정하였다. 과육경도는 경도계(KM-5, Fujiwara, Tokyo, Japan)를 이용하여 측정하였다. 10개의 과실의 특성에 대한 평균을 1반복으로 하여 총 3반복으로 조사하였다.
대추 유전자원 당 30개의 핵을 이용하여 핵의 종경, 횡경, 무게 및 핵형지수를 조사하였다. 종자 형성 유무는 핵 특성 조사 후 고무망치를 이용하여 핵을 갈라내어 충실한 배와 배유를 함유한 종자가 형성되었는지 조사하였다. 10개 핵의 특성에 대한 평균을 1반복으로 하여 총 3반복으로 조사하였다.
기능성 성분 분석
10월에 수확한 대추 과실의 핵을 제거한 과피와 과육을 동결건조기(PVTFD-50R, Ilshin Lab Co. LTd., Busan, Korea)를 이용하여 건조하였다. 건조된 시료는 분쇄기(1093 Cyclotec Mill Unit, Foss, Hoganas, Sweden)로 분쇄하였다. 건조시료 2 g에 18 mL의 증류수를 첨가하여 상온에서 2시간 동안 진탕 배양하였다. 이 후, 4℃에서 8,000 rpm으로 20분 동안 원심분리 후 12 mL의 상층액을 수집하였다. 수집한 상층액은 감압여과(Adventec No.2, Adventec, Tokyo, Japan)한 후 3반복으로 분석에 이용하였다.
총 폴리페놀 함량 분석은 Dewanto et al. (2002)의 방법으로 측정하였다. 추출시료 0.5 mL에 2% Na2CO3용액 1 mL를 첨가 후 3분간 상온에 방치시켰다. 이 후, 50% 2 N Folin-Ciocalteureagent (Sigma-Aldrich Co., St. Louis, Mo, USA) 0.5 mL를 첨가 후 상온에 30분 동안 방치하였다. Spectrophotometer (Cary 100 UV/Vis., Agilent Tec., Santa Clara, USA)를 이용하여 반응액의 흡광도를 750 ㎚에서 측정하였다. 표준물질은 gallic acid를 사용하여 검량선을 작성하였고, 총 폴리페놀 함량은 추출물 중의 mg gallic acid equivalent (GAE, dry basis)로 나타내었다.
전자공여능(electron donating ability, EDA)은 Blois (1958)의 방법을 변형하여 분석하였다. 추출물 0.2 mL에 0.4 mM 1,1-diphenyl-2-picrylhydrazyl (DPPH)용액 0.8 mL를 첨가한 후 상온에서 10분간 방치한 다음 525 ㎚에서 흡광도를 측정하였다. 유전자원 당 3반복으로 조사하였다. 전자공여능은 시료 첨가구와 비첨가구의 흡광도 차이로 다음의 식에 의하여 계산되었다.
DPPH radical scavenging activity (%) = {1 - (Absorbance of sample at 525 ㎚ / Absorbance of control at 525 ㎚)} × 100
통계분석
주요 특성의 통계 처리는 SPSS (statistical package for social science, version 24.0, SPSS Inc., Chicago, IL USA)프로그램을 이용하여 5% 유의수준에서 던컨의 다중검정방법(Duncan’s multiple range test)을 수행하였다.
주성분분석(principal component analysis) 또한 SPSS 프로그램을 이용하여 고유값, 주성분의 기여도, 주성분내 각 형질의 기여도를 산출하였다. 주성분분석으로 계산된 형태형질의 측정값을 Z점수로 표준화하여 계층적 군집분석을 수행하였다.
결과
생육 특성
‘일본’을 포함한 10개 대추 유전자원을 대상으로 발아기, 개화기, 숙기 및 개화성을 조사한 결과, ‘일본’의 발아기는 4월 15일로 다른 유전자원들의 발아기와 유사하였으나, ‘국광’만 4월 19일로 가장 늦게 발아하였다(Table 2). 개화기는 ‘일본’과 ‘호대추’, ‘국광’, ‘무등’, ‘상왕’은 6월 19일로 가장 늦었으며 ‘보은’과 ‘복조는’ 6월 13일로 가장 빨랐다. 대추의 숙기는 10월 상순으로, ‘일본’은 숙기가 10월 9일이었으며, ‘보은’, ‘상왕’, ‘티벳왕’과 같았다. ‘국광’의 숙기는 10월 12일로 다른 유전자원들과 비교하여 늦은 경향을 보였다. 대추나무의 특이한 생육 특성인 개화성을 조사한 결과, ‘일본’은 13시부터 꽃받침이 열리기 시작하여 14시부터 15시에 개화하는 오후 개화성이었다. 대부분의 유전자원들 또한 오후 개화성이었으나 ‘보은’만 9시부터 10시 사이에 꽃받침이 열리기 시작하여 오전 개화성으로 조사되었다.
Table 2.
Growth characteristics of bud burst date, flowering date, harvesting date and blooming type among 10 jujube accessions
엽 특성
대추 유전자원의 엽 형질 조사 결과(Table 3), ‘일본’의 엽폭과 엽장, 엽병장은 각각 2.8, 5.5, 5.0 ㎝로 다른 9개 유전자원들과 비교하여 유의적으로 가장 짧았다. 엽형지수는 ‘일본’이 2.0으로 가장 높았으며 모든 유전자원들이 1.6 이상의 엽형지수를 보여 타원형의 엽형을 나타내었다. 10개 대추 유전자원의 엽 거치형태는 둔거치와 예거치로 구분되었으며, ‘일본’, ‘추석’, ‘국광’, ‘무등’, ‘상왕’은 예거치로 조사되었다. 엽저는 ‘일본’, ‘보은’, ‘국광’, ‘무등’, ‘상왕’은 예저로 분류되었고, ‘티벳왕’은 심장저로, 나머지 4개 유전자원은 원저로 분류되었다.
Table 3.
Leaf characteristics among 10 jujube accessions
과실 특성
대추 유전자원의 7가지 과실특성을 조사한 결과, ‘일본’의 과실의 크기는 횡경과 종경이 각각 23.8, 27.1 ㎜로 가장 작았으며, ‘국광’은 횡경과 종경이 40.5, 44.0 ㎜로 가장 컸다(Table 4, Fig. 1). 과중은 과실 횡경과 종경이 가장 짧은 ‘일본’이 7.8 g으로 가벼웠으며, 과실 횡경과 종경에 비례하여 과중이 무거웠다. ‘일본’의 당도는 26.0 °Brix로 한국 유전자원인 ‘추석’(33.6 °Brix)에 비해 낮았으나, 과실의 경도는 3.4 ㎏∙5 ㎜Ø-1로 높았으며 통계적으로 유의성이 있었다. 10개 대추 유전자원들의 과형지수는 최소 1.1에서 최대 1.5로 1.1의 과형지수를 나타낸 ‘일본’, ‘국광’, ‘티벳왕’은 편주형이었다. ‘복조’, ‘추석’, ‘금성’, ‘호대추’, ‘무등’은 원주형이었으며, ‘보은’은 난원형, ‘상왕’은 장원형으로 조사되었다.
Table 4.
Fruit characteristics of among 10 jujube accessions

핵 특성
핵의 종경과 횡경, 무게는 ‘일본’과 ‘보은’을 제외한 8개 유전자원은 과실 크기에 비례하였다(Table 5) ‘일본’의 핵의 횡경과 종경은 각각 8.8, 15.4 ㎜로 조사되었으며, 무게는 0.6 g으로 과실에 비해 큰 편이었다. 핵형지수는 10개 유전자원에서 최소 1.8 (‘일본’)에서 최대 3.7 (‘무등’)의 범위를 나타내어 모두 타원형 또는 방추형의 핵형을 나타냈다. ‘일본’, ‘보은’, ‘추석’, ‘국광’, ‘티벳왕’은 타원형의 핵형을, 나머지 5개 유전자원은 방추형의 핵형으로 조사되었다. 교배육종을 통한 품종 육성 시 가장 중요한 요인인 종자 형성 유무를 조사한 결과, ‘복조’, ‘추석’을 제외한 8개 유전자원들은 종자가 형성되었다. ‘일본’의 경우 충실한 종자 형성율이 50% 이상으로 높았다(Data not shown).
Table 5.
Fruit stone characteristics among 10 jujube accessions
과실의 기능성
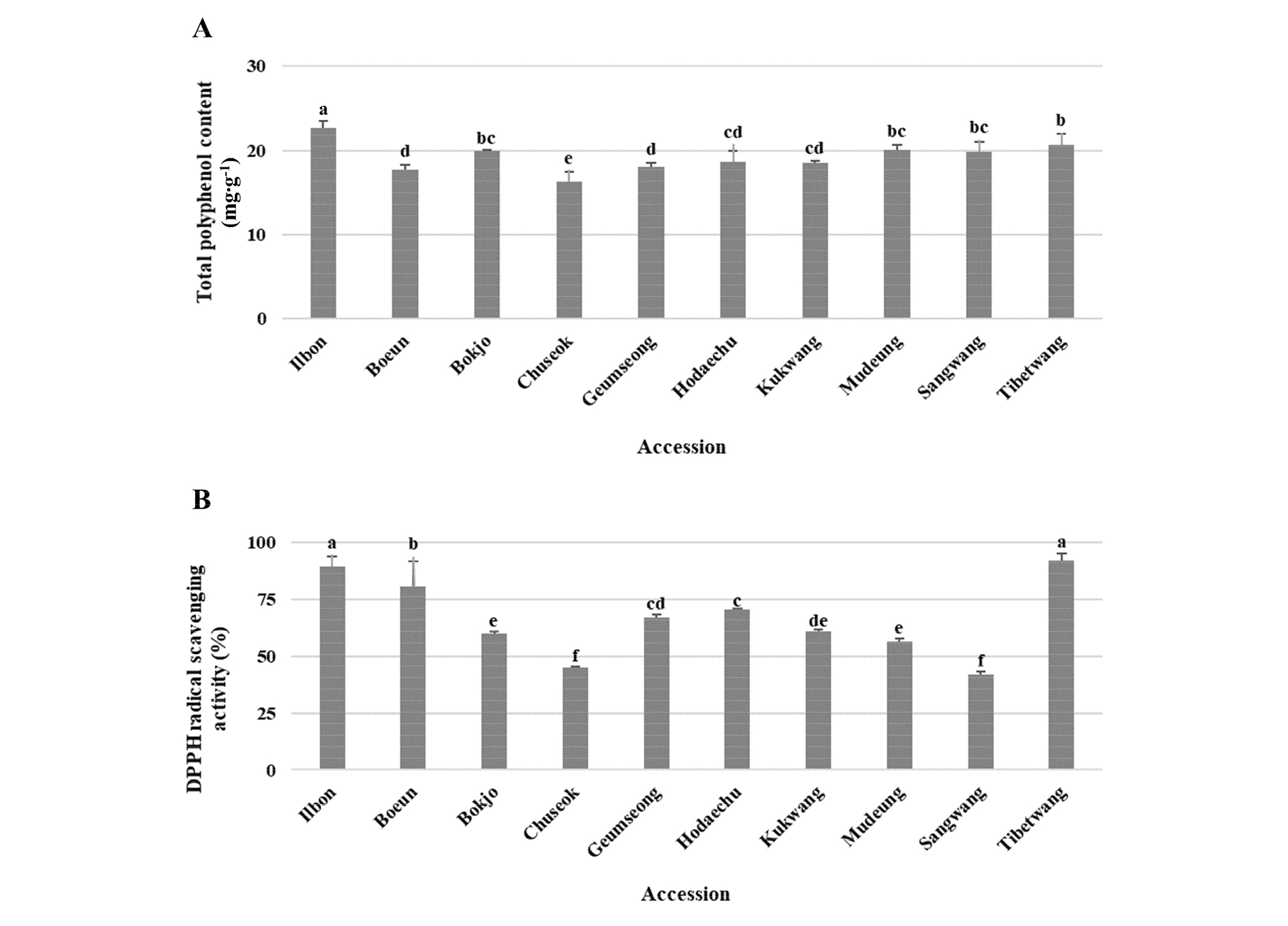
대추 유전자원들의 과실에 포함된 총 폴리페놀 함량과 DPPH radical 소거능을 분석한 결과(Fig. 2), 총 폴리페놀 함량은 ‘일본’이 22.65 ㎎∙g-1으로 가장 높았고 ‘추석’이 16.25 ㎎∙g-1으로 가장 낮았다. 총 폴리페놀 함량이 높은 유전자원들이 DPPH radical 소거능 또한 높았으며, ‘일본’ 다음으로 총 폴리페놀 함량이 높았던 ‘티벳왕’(20.69 ㎎∙g-1)은 DPPH radical 소거능이 92.23%로 가장 높았다. ‘일본’의 DPPH radical 소거능은 89.44%로 조사되었으나 ‘티벳왕’과 유의적인 차이가 없었다.
주성분 및 군집 분석
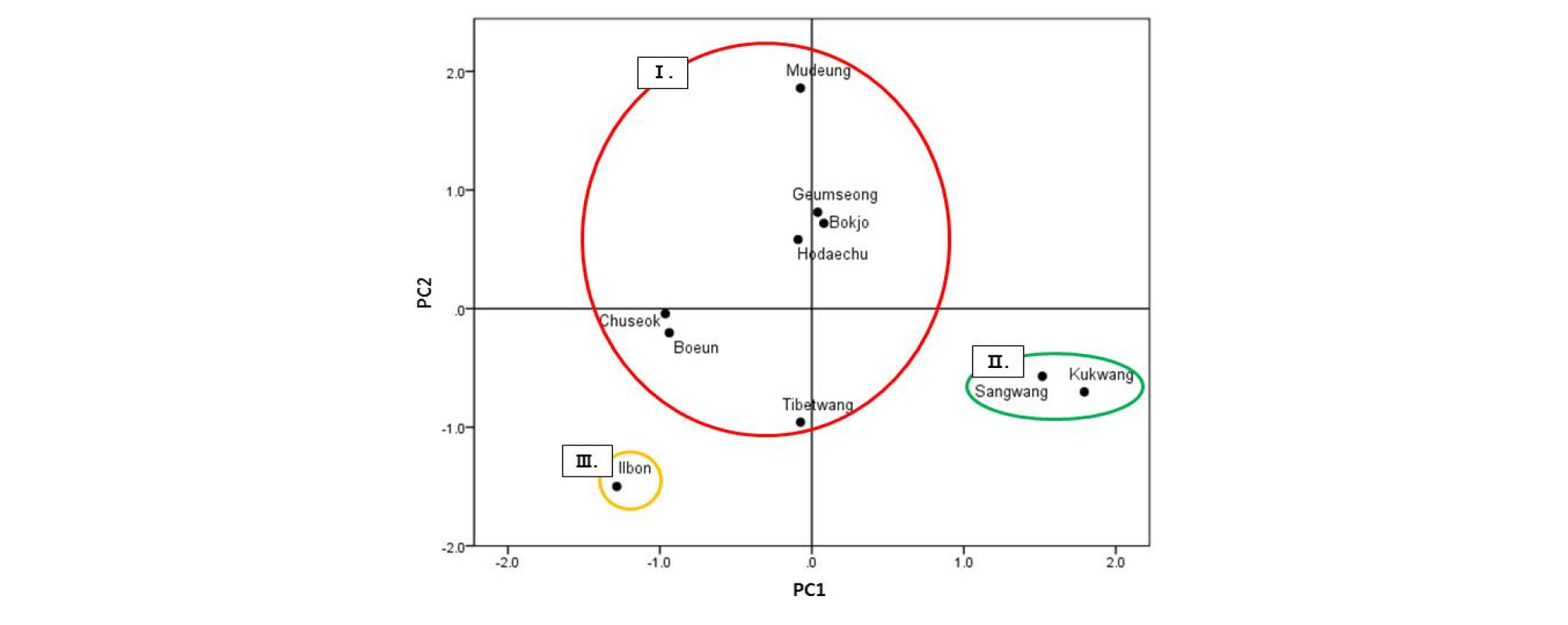
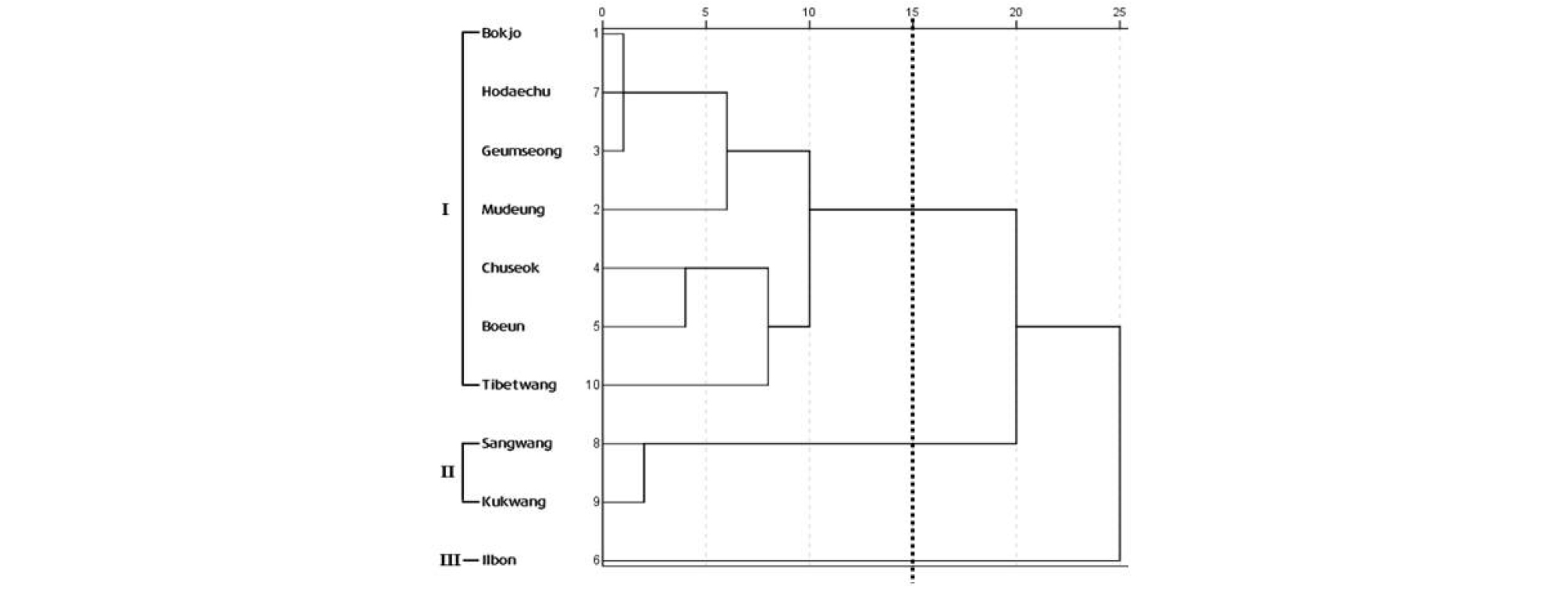
‘일본’을 포함한 주요 대추 유전자원 10개의 형태형질 중 14개의 양적형질을 이용하여 주성분분석을 수행한 결과, 3개의 군집으로 구분되었다(Fig. 3). 제1주성분은 엽폭(0.954), 엽장(0.945), 엽병장(0.936), 엽형지수(0.907), 과실횡경(-0.837), 과실종경(0.792) 등 엽과 과실 크기 관련 특성이었으며, 제2주성분은 과형지수(0.963), 과중(0.948) 과실 당도(0.929), 과육경도(-0.820), 핵횡경(0.787), 핵종경(0.734), 핵형지수(0.569)로 과실과 핵의 크기 관련 특성이었다. 제3주성분은 핵 무게(-0.947)였다. 제1, 2, 3주성분은 각각 39.41, 39.15, 12.87%의 eigenvalue를 나타내어 전체 분산에 대해 78.56%를 설명할 수 있는 제2주성분 수준에서 유전자원들을 분류하였다. 1번 군집은 한국 유전자원인 ‘보은’, ‘복조’, ‘추석’, ‘금성’, ‘무등’과 중국 유전자원인 ‘호대추’와 ‘티벳왕’으로 구성되었다. 중국 유전자원인 ‘국광’과 ‘상왕’은 2번 군집으로 구성되었으며, ‘일본’은 9개 유전자원들과 뚜렷하게 구분되어 3번 군집을 형성하였다. 주성분분석 결과와 동일하게 계층적 군집분석 결과는 유사성 15수준에서 3개의 군집으로 분리되었다(Fig. 4). 각 군집에 속한 유전자원은 주성분분석 결과와 동일하였다.
고찰
대추는 꽃이 작아 인공수분이 어려우며 단위결과가 용이하고 충실한 종자가 형성되지 않아 교배육종이 어렵다(Ackerman, 1961). 충청북도농업기술원 대추연구소에서 보유하고 있는 대추 유전자원 중 ‘일본’의 형태적 특성이 다른 유전자원들과 비교하여 육안상 뚜렷한 차이를 나타내고, 충실한 종자가 발달하는 것으로 관찰되어 추후 유전 연구를 위한 육종 소재로의 이용 가능성을 확인하고자 주요 대추 유전자원들과 형태형질을 비교하고 유연관계를 확인하였다.
대추 10개 유전자원의 발아기는 4월 15일부터 19일 사이로 조사되어 유전자원 간의 저온요구도의 차이가 크지 않은 것으로 판단된다. 또한 개화기의 경우, ‘일본’은 6월 19일로, ‘호대추’, ‘국광’, ‘무등’, ‘상왕’ 등의 유전자원과 개화기가 동일하고 모두 오후 개화성으로 조사되어(Table 2) ‘일본’을 이용한 교배육종 시 교배 모본 또는 부본으로 이용 가능한 유전자원들이다.
엽과 과실, 핵의 형태형질을 조사한 결과, ‘일본’의 엽 형질과 과실특성은 다른 9개 유전자원들과 비교하여 뚜렷한 크기의 차이를 나타냈다(Tables 3 and 4, Fig. 1). ‘일본’의 엽폭과 엽장, 과실 종경과 횡경 모두 9개 유전자원들과 비교하여 유의적으로 짧았다. 과실 크기에 비례하여 과중 또한 ‘일본’이 가장 작았다. ‘일본’의 과실 당도는 ‘국광’, ‘상왕’, ‘티벳왕’과 함께 유의적으로 낮아 추후 당도나 과실 크기 증진을 위한 품종 육성 시 ‘추석’ 및 ‘호대추’ 등과의 교배를 통해 품질향상이 가능할 것으로 생각된다.
Z. jujuba에 속하는 대추 유전자원들 중에는 종자가 발달되지 않는 개체들이 다수 존재하여(Asatryan and Tel-Zur, 2013; Tian and Ma, 1987) 교배육종을 통한 품종육성은 아직까지 보고된 바 없다. 본 연구에서 이용된 10개 대추 유전자원 중 ‘복조’와 ‘추석’은 핵은 존재하나 종자가 퇴화되어 교배육종에 활용하는 것이 거의 불가능하다. 종자가 발달한 나머지 8개 유전자원 중 ‘일본’은 정상적인 종자 형성율이 50% 이상으로 높았으며, ‘보은’은 약 20%로 조사되었고, 나머지 6개 유전자원들은 5% 미만으로 정상적인 배와 배유를 포함한 충실한 종자의 비율이 매우 낮았다. 그러므로 ‘일본’은 다른 유전자원들과 비교하여 정상적인 종자 형성율이 매우 높아 추후 교배를 통한 실생 획득에 이용 가능하며, 세대진전을 통한 대추의 유전연구를 가능하게 할 수 있는 유용한 유전자원이다.
또한 ‘일본’은 기능성 성분인 총 폴리페놀 함량과 DPPH radical 소거능이 다른 유전자원과 비교하여 유의적으로 높았다(Fig. 2). 특히 ‘일본’은 국내 주요 재배 품종인 ‘무등’, ‘금성’, ‘복조’, ‘보은’(Park and Kim, 2016) 보다 항산화 능력이 높아 추후 고기능성 대추 품종 육성을 위한 육종 소재로 활용가치가 높다.
엽 형질과 과실특성은 양적형질로 환경의 영향을 받기 쉬우나 유전자원들의 형태형질 특성 데이터를 이용하여 유전자원 간의 유연관계를 구명할 수 있다(Assefa et al., 2020; Jang et al., 2006; Kim et al., 2013; Kim et al., 2017). 특히 Jang et al. (2006)은 대추 유전자원의 형태형질을 이용하여 품종 간의 유연관계를 밝혔으나, 현재까지 ‘일본’의 유연관계는 확인되지 않아 본 연구를 통해 조사된 형태형질 14항목을 이용하여 주성분분석을 수행하였다.
엽과 과실 크기가 뚜렷하게 작은 ‘일본’은(Tables 3 and 4) 9개 유전자원들과 비교하여 형태적 다형성이 커 새로운 분류군을 형성하였다(Figs. 3 and 4). 주성분분석 및 유연관계 분석 결과에서 동일하게 1번 군집은 ‘호대추’, ‘티벳왕’을 제외하고 한국 원산의 유전자원들로 구성되었으며, 2번과 3번 군집은 각각 중국과 일본 원산의 유전자원들로 분류된 것으로 보아 대추 유전자원들은 지리적 기원에 따른 다양성을 나타내는 것으로 생각된다.
세계적으로 보존 중인 대추 유전자원들은 종자 형성능이 낮아 교배육종을 통한 품종육성 사례가 전무하다. 형태적 다양성이 높은 ‘일본’은 충실한 종자 형성능과 발아율을 보이며 기능성 성분이 높아 추후 교배육종을 통해 기능성 성분이 높은 대추 품종을 육성할 수 있을 것으로 전망된다. 또한 본 연구를 통해 조사된 대추 유전자원들의 형태형질은 교배육종의 소재를 선정하는 데에 기여할 것이다.
적요
본 연구는 충청북도농업기술원 대추연구소에서 보유 중인 대추 유전자원 중 형태적 다형성을 나타내는 ‘일본’과 주요 대추 유전자원의 형태형질을 조사하고, 대추 육종 소재로서 ‘일본’의 가능성을 확인하기 위해 수행되었다. ‘일본’은 주요 대추 유전자원과 비교하여 엽과 과실의 크기가 뚜렷하게 작고 당도가 낮았다. 하지만 과실에 폴리페놀 함량이 가장 높았으며, 항산화 능력 또한 가장 높았다. 또한 ‘일본’은 종자형성능이 가장 높았다. 대추 유전자원들의 형태형질을 이용하여 분석한 결과, 지리적 기원에 따라 유전자원들이 분류되었다. 특히 ‘일본’은 한국 및 중국 유전자원들과 명확히 구분되었다. 따라서 종자형성능이 우수하고 기능성 성분이 높은 ‘일본’은 추후 대추 유전연구 및 교배육종을 통한 품종 육성에 유용하게 이용될 수 있을 것으로 생각된다.
