

서언
분류군의 기재
Ludwigia peploides subsp. montevidensis (Spreng.) P.H. Raven, Reinwardtia 6: 395 (1964)
한국산 여뀌바늘속과 근연종의 검색표
고찰
적요
서언
침입외래식물은 의도적 또는 비의도적으로 도입되어 전 세계적으로 확산되고 있으며, 자생종의 감소나 절멸을 초래하는 등 생물다양성에 부정적인 영향을 미치는 것으로 알려져 있다(Mooney, 2001). 이에 국내에서도 침입외래식물의 작물 또는 축산업에 대한 부정적 영향(Ryu et al., 2018), 하천 하안지역의 교란 등을 우려한 연구와 관리방안(Lee et al., 2015; Lim et al., 2009)에 대한 연구들이 진행되고 있다. 침입력이 강한 외래식물은 자생식물 보다 경쟁력 있는 생존전략을 가지고 있으며, 생태 적응력이 뛰어나고 무성생식을 하며 성장이 빠르고 번식력과 발아력, 그리고 생식시스템의 적응성이 높은 특성을 보이는데(Pysek et al., 1995), 바늘꽃과(Onagraceae Juss.)의 여뀌바늘속(Ludwigia L.) 식물들은 이런 특징들을 대부분 보유하고 있다. 또한 육지와 물 모두에서 자라며, 개체 확산이 매우 빨라 가장 공격적인 외래 식물200종 안에 포함되기도 하였다(Cronk and Fuller, 2014).
여뀌바늘속은 바늘꽃과 식물 중 가장 크고 다양한 분류군들을 포함하고 있으며, 전 세계적으로 약 82여종이 온대와 열대 지역의 습지와 산지 등 다양한 지역에 넓게 분포하는 수륙양육의 분류군이다(Eyde, 1977; Zardini et al., 1991a). 이들은 형태학적으로 매우 유사하고 영양기관의 다양한 변이로 인하여 종 동정이 매우 어려운 분류군 중 하나로 알려져 있는데, 이는 대부분의 종이 지리적으로 넓게 분포하고 자생지의 생육환경에 따라 표현형이 매우 다양하게 나타나기 때문이다(Zardini et al., 1991a). 한편 국내 여뀌바늘속 식물은 여뀌바늘(L. prostrata Maxim.)과 눈여뀌바늘(L. ovalis Miq.) 2종이 알려져 있으며(Lee, 1980; Park, 2007), 도감상의 단순 기재만 있을 뿐 이들의 분류학적 연구는 전무하다.
본 연구에서는 수원시 하천변에서 발견된 여뀌바늘속의 Ludwigia peploides subsp. montevidensis (Spreng.) P.H. Raven에 대한 형태학적 기재, 생태 사진, 주요 형질에 대한 도해뿐 만 아니라 한국산 여뀌바늘속 식물의 종 동정을 위한 명확한 검색표를 제시하여 향후 국내 침입외래식물 관리를 위한 기초자료를 제공하고자 한다.
분류군의 기재
Ludwigia peploides subsp. montevidensis (Spreng.) P.H. Raven, Reinwardtia 6: 395 (1964)
Jussiaea montevidensis Spreng., Syst. Veg., ed. 16 [Sprengel] 2: 232 (1825)
Ludwigia adscendens var. montevidensis (Spreng.) H. Hara, J. Jap. Bot. 28: 291 (1953)
Ludwigia peploides var. montevidensis (Spreng.) Shinners, Sida 1: 386 (1964)
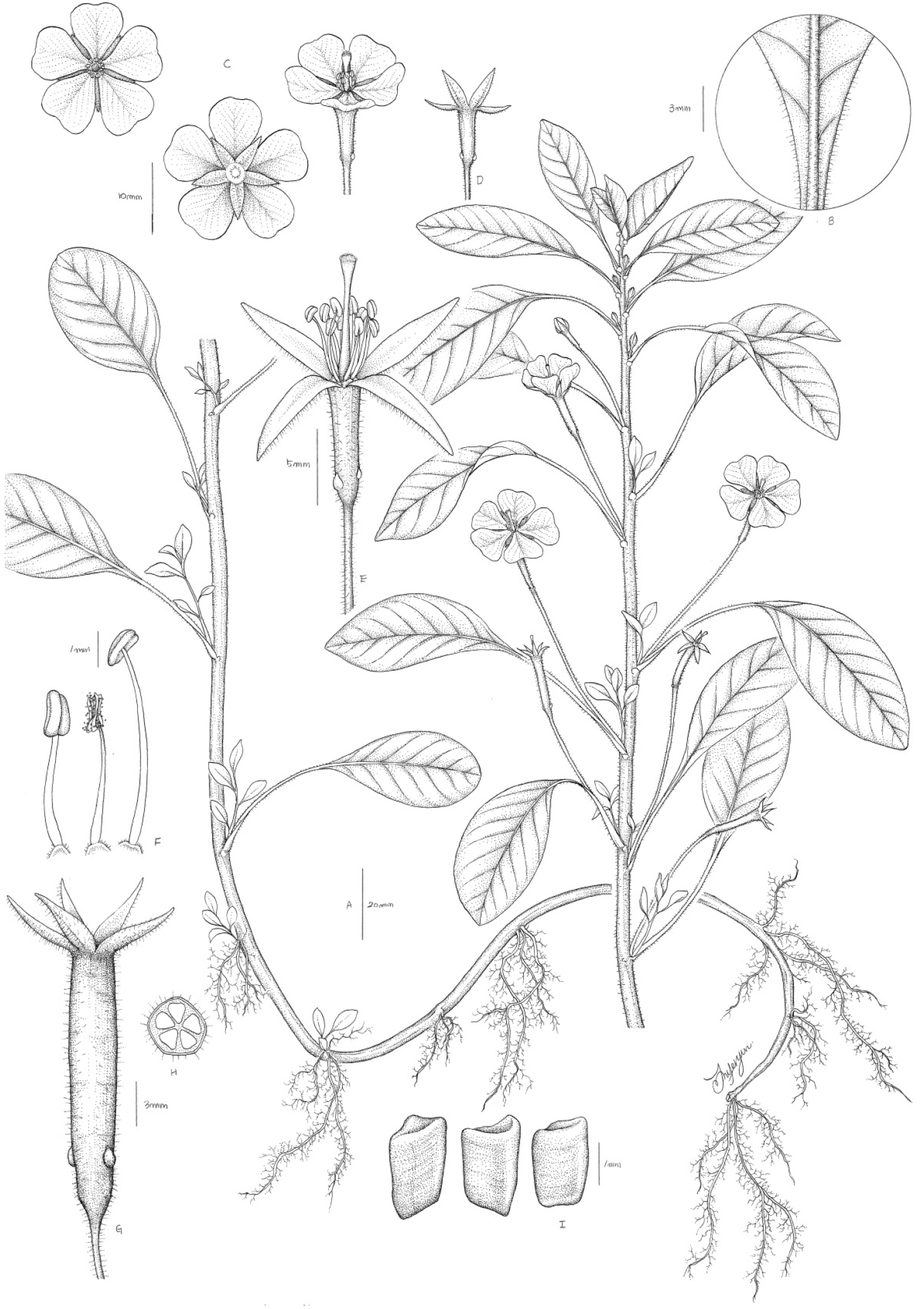
다년초이다. 줄기는 넓게 뻗어 나가며 마디에서 뿌리를 내리거나 물에 떠 있으며, 연모 또는 끈적한 물질을 분비하는 털로 덮여 있다. 물에 떠 있는 줄기는 털이 없기도 하다. 잎은 어긋나며 잎자루는 길이 2.0-5.5 ㎝이다. 잎몸은 길이 3.5-8(10) ㎝의 긴 타원형이고 거치는 없으며, 기부는 좁은 쐐기모양이고, 선단부는 예두, 예첨두, 둔두이며, 잎맥은 7-14개이다. 물에 뜨는 잎은 어긋나며 길이 2.0-6(8) ㎝로 원형, 타원형 또는 넓은 타원형이고, 선단부는 둥글며, 잎자루는 길이 1.5-5.5 ㎝이다. 꽃은 위쪽 줄기의 잎 겨드랑이에 한 개씩 달리며, 꽃자루는 길이 1.5- 4.8 ㎝이고 털이 많다. 꽃받침은 5장으로 길이 5-12 ㎜이며 피침형 또는 좁은 삼각형에서 넓은 삼각형으로 털이 밀생하거나 없기도 하다. 꽃잎은 5장으로 도란형이고 길이 1.2-1.7 ㎝이며 밝고 진한 노란색이고 가운데 부분은 바깥쪽보다 색이 진한 편이다. 수술은 10개이고 길이 2.5-5 ㎜이며, 수술대는 밝은 노란색이고 꽃부리에 붙은 것들은 다소 짧기도 하다. 암술은 1개이고, 암술대는 노란색이며 아래쪽 반 이상이 긴 털로 덮여 있다. 암술머리는 길이 약 1 ㎜ 정도의 깊이로 5열 한다. 열매는 삭과로 길이 1.5-3 ㎝이며 밝은 갈색이고 실린더형이며 5개로 약간 각이 져 있고 짧은 털이 있다. 포는 열매 아래쪽 또는 중간 정도에 위치하며 삼각형 모양이고 길이는 약 1 ㎜이다. 종자는 길이 1-1.3 ㎜로 각 심피마다 1열 배열한다.
국명: 꽃여뀌바늘(Kkot-yeo-kkwi-ba-neul)
개화기: 5~10월
결실기: 6~11월
관찰표본: Seohocheon (river), Jeongja-dong, Jangan-gu, Suwon-si, Gyunggi-do, 6 Aug 2014, YSM14041 (KH)
논의: 본 종은 Sprengel (1825)에 의해 우루과이 몬테비데오(Montevideo)지역에서 채집된 표본(Sello s.n.)에 근거하여 Jussiaea속(Jussiaea L.)의 신종으로 기재되었고, 이후 여러 학자들에 의해 여뀌바늘속으로 편입되어 분류되었으며(Wagner, 2007), 최근 Raven (1964)은 전 세계산 Ludwigia속의 연구에서 본 종을 L. peploides (Kunth) P.H. Raven의 아종으로 분류학적 처리를 수행한 바 있다. 이와 반대로 일부 학자들은 본 종을 독립된 종으로 인정하지 않고 모종인 L. peploides로 통합하여 인식하기도 하였지만(Kim et al., 2018), L. peploides는 식물체 전체에 털이 없고 매끈하며 열매(capsule) 기부에 포(bracteoles)가 위치하는데 반해, L. peploides subsp. montevidensis는 식물 전체에 털이 있고 열매의 기부에서 1/5 정도 윗부분에 포가 위치하는 특징으로 뚜렷이 구분된다(Jiarui et al., 2007; Raven, 1963; Zotos et al., 2006) (Fig. 1-G). 따라서 Kim et al. (2018)이 언급한 L. peploides (물여뀌바늘)는 그 기재와 수록된 식물 사진들로 보아 L. peploides subsp. montevidensis의 오동정으로 판단된다. 또한 꽃여뀌바늘은 꽃자루가 길며 꽃받침이 5장이고, 꽃의 직경이 3-3.5 ㎝ 정도로 크며, 선명한 노란색 꽃잎 5장을 가지고 수술의 수가 꽃받침 수의 두 배인 특징을 가지므로(Fig. 2-C, D, E) 꽃자루가 없고 꽃받침이 4장이며, 꽃의 직경이 1 cm 정도로 보다 작거나 아예 꽃잎이 없고 수술 수가 꽃받침 수와 같은 국내 분류군들[여뀌바늘(L. prostrata), 눈여뀌바늘(L. ovalis)]과 뚜렷이 구분된다. 본 종의 국명과 관련하여 국내에서는 ‘물앵초’, ‘물달맞이꽃’, ‘물여뀌바늘’ 등으로 불리우고 있으나(Kim et al., 2018), 본 종은 바늘꽃과의 여뀌바늘속 식물로서 앵초속 및 달맞이꽃속 식물과 관련이 없고, ‘물여뀌바늘’이라는 이름은 원예종인 Ludwigia repens J.R. Forst.의 국명으로(Korea National Arboretum, 2010) 이미 알려져 있기 때문에 상기 국명들은 본 종을 인식하는데 있어 혼동을 초래할 수 있다. 따라서 본 연구에서는 본 종이 원예종으로 식재될 만큼 노란색의 큰 꽃을 가지고 있으며 개화기가 긴 특징에 근거하여 ‘꽃여뀌바늘’로 국명을 개칭하였다.

Fig. 1.
Photographs of L. peploides subsp. montevidensis (Spreng.) P.H. Raven A. Lake Seoho (Suwon-si), B. Lake Mansuk (Suwon-si) in late October, C. Stems, D. Floating leaves, E. Flower bud, F. Flower, G. Capsule (arrow: bracteoles), H. Viscid hairs (arrow: sticky liquid). Photographs by Hye-Won Kim, Chang-Seok Jang and Soo Hyun Park.
한국산 여뀌바늘속과 근연종의 검색표
1. 꽃은 4수성이다; 꽃의 직경은 1 ㎝ 미만으로 작거나 없으며 꽃자루가 없다. 수술의 수는 꽃받침의 수와 같다.
2. 일년생이다. 꽃잎은 노란색이며 길이 0.5 ㎝ 정도로 작아 눈에 잘 띄지 않는다. 줄기는 곧게 자란다
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・L. prostrata Roxb. 여뀌바늘
2. 다년생이다. 꽃잎은 없다. 줄기는 기어 자라며 마디에서 뿌리를 내린다.・・・・・・・・・・・・・・・・・・・・・L. ovalis Miq. 눈여뀌바늘
1. 꽃은 5수성이다; 꽃의 직경은 3-3.5 ㎝ 정도로 크고 노란색으로 눈에 잘 띄며 꽃자루가 1.5-4.8 ㎝ 정도로 긴 편이다; 수술의 수는 꽃받침 수의 두 배이다.
3. 꽃이 피는 줄기는 주로 매끈하며 가끔 약간 털이 있기도 하나 끈적이지는 않는다. 포는 삭과의 기부에 위치한다.・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・L. peploides subsp. peploides
3. 꽃이 피는 줄기는 끈적이는 물질을 내는 긴 털로 덮여 있으나 드물게 털이 없기도 하다. 포는 삭과의 기부에서 1/5정도 위쪽 부분에 위치한다.・・・・・・・・・・・・・・・・・・・・・・・・・・・・・L. peploides subsp. montevidensis (Spreng.) P.H.Raven 꽃여뀌바늘
고찰
습지나 저수지, 하안 생육지의 경우 수위변동에 의한 스트레스와 교란 및 인간에 의한 교란이 빈번하게 발생하므로 외래식물의 유입과 확산이 용이한 편이다(Cho and Cho, 2015). 물가에 자라는 꽃여뀌바늘은 남아메리카가 원산지로 알려져 있으며 현재 미국을 비롯하여 호주, 뉴질랜드, 북유럽(프랑스, 벨기에) 및 유럽 남부의 그리스까지 전 세계적으로 퍼져 있고(Zotos et al., 2006) 가장 침입력 강한 수생식물 중 하나로 평가를 받아 생태계교란위험종으로서 관리되고 있다(EPPO, 2011). 특히 포복성의 줄기는 각 마디에서 뿌리가 생성되며 줄기가 잘릴 경우에도 육상뿐 만 아니라 수중에서도 생육이 가능하기 때문에 본 종이 확산되는 주요 원인으로서 지속적인 관리가 필요한 부분이기도 하다(EPPO, 2011). 또한 뿌리는 물속에 가라앉거나 떠 있는 줄기들을 개체군 매트로서 고정시키고 새로운 줄기가 자랄 수 있도록 지지해주는 역할을 하며, 물에 떠 있는 잎 또한 단단한 개체군 매트를 형성하여 점차 확산되면서 물 표면을 빠르게 뒤덮어 나가고 호수, 작은 못이나 수로, 좁은 하천 등의 물 흐름을 제한하여 침전물을 가둬 두는 등 수질의 악화를 야기 시킬 수 있다(Zardini et al., 1991b; Zotos et al., 2006; Thiébaut and Dutartre, 2012). 게다가 물에 뜨는 줄기로 수면을 완전히 덮고, 거대 군락을 형성함에 따라 어류나 다른 식물의 생존을 어렵게 하여 수생태계에 부정적인 영향을 줄 수 있다(Stroh, 2006; Lee et al., 2015). 생식에 있어서도 여뀌바늘속 식물은 일반적으로 충매화이지만 꽃여뀌바늘은 2배체(2n=16) 식물로서 자가수분이 가능하고 무성생식을 통한 번식도 활발하며(Dandelot et al., 2005; Zotos et al., 2006; Ruaux et al., 2009) 종자생산력이 매우 높기 때문에 빠른 속도의 확산이 용이하다.
2015년도부터 시행된 유럽연합(EU)의 외래종에 대한 규정(IAS Regulation)에서는 L. peploides를 위험 식물로 지정하고 수입이나, 보유, 판매, 재배 등을 제한하고 있다(European Union, 2017). 수원시에서는 꽃여뀌바늘이 최초 발견된 서호천뿐 아니라 상류지역인 만석거에서도 큰 집단을 찾았는데 하천의 흐름을 고려할 때 만석거에 식재된 개체가 관리 미흡으로 하류인 서호천으로 퍼져나간 것으로 판단된다. 이런 개체들은 하천을 따라 개체수가 매년 증가하고 있으며, 동절기에도 큰 집단의 흔적들을 쉽게 관찰할 수 있다(Fig. 1-A, B). 또한 수원 이외에도 양평, 오산, 창녕 등지에서도 개체들의 기록을 확인할 수 있었는데 이들 또한 식재로 시작해 야생으로 퍼진 것으로 추정된다. 외국의 정책이나 꽃여뀌바늘의 생태적 특성들을 고려해 볼 때, 꽃여뀌바늘은 생태계교란종으로서 확산될 우려가 있으므로 식재가 아닌 지역에서 발견될 경우뿐 아니라 식재 지역에서도 지속적인 관심을 가지고 모니터링을 하며 개체군이 더 퍼지지 않도록 관리 방안을 신속히 강구하여야 할 것이다.
적요
의도적 도입 또는 비의도적 도입에 의해 국내에 확산되고 있는 침입외래식물은 경제적, 환경적인 해를 가할 수 있으며 자생종의 감소나 절멸을 초래하는 등 생물다양성에 부정적인 영향을 미치는 것으로 알려져 있다. 특히 습지나 저수지 및 하안 생육지는 스트레스와 교란에 의해 외래식물 유입과 확산이 용이하므로 국립수목원에서는 이를 관리하기 위한 연구를 지속적으로 수행하고 있다. 본 연구에서는 수원시 하천변에서 발견된 바늘꽃과에 속하는 L. peploides subsp. montevidensis (Spreng.) P.H. Raven가 생태계교란종으로 확산될 우려가 있어 본 종에 대한 기재, 도해, 칼라도판 등을 첨부하여 침입외래식물 관리를 위한 기초자료를 확보하고자 한다.
