

Introduction
Materials and methods
Plant material
Morphological characteristics
Phenological characteristics
Fruit characteristics
Data analysis
Results and discussion
Correlations between traits
Introduction
Plums, which are grown in temperate zones, rank fifth among deciduous fruits produced in the world (FAO, 2016). In the Republic of Korea, plums rank sixth in place among deciduous fruits in production, following apple, persimmon, peach, grape, and pear (KOSIS, 2016). In 2016, the production and cultivated area of plums were 78,857 metric tons and 7,033 ha, respectively, in the Republic of Korea (KOSIS, 2016). Plums belong to the family Rosaceae, which consists of approximately 3,000 fruit species and 88-100 genera (Xiang et al., 2016). Plum and other stone fruits, such as apricot, cherry, and peach, belong to the genus Prunus (Byrne, 2012), which includes several subgenera, such as Amygdalus, Cerasus, and Prunophora (Topp et al., 2012). Almond and peach belong to the subgenus Amygdalus, whereas apricot and plum belong to the subgenus Prunophora. Plums are distributed among 19-40 species, with more than 6,000 varieties (Hedrick, 1911; Blažek, 2007), where the diversity of plum is the largest in Prunus. Therefore, plums are considered as a link to other subgenera in Prunus and as the center of the Prunus genetic stage (Watkins, 1976).
There are three major plum groups according to their species: European, American, and Asian (Topp et al., 2012). The major species of the European plum is Prunus domestica, which originated from the south of the Caucasus Mountains through to the Caspian Sea. P. domestica is a hexaploid (2n = 6x = 48) and is generally thought to have resulted from a cross between the diploid Prunus cerasifera (2n = 2x = 16) and the tetraploid Prunus spinosa (2n = 4x = 32), which also belong to the European plum group (Faust and Surányi, 1999). P. cerasifera, known as the cherry plum or myrobalan plum, has been widely used as a rootstock and is found in the parentage of many modern Asian plums (Boonprakob et al., 2001). P. spinosa is known as the blackthorn or sloe, and its fruits are generally small, sour, and astringent (Buttner, 2001).
Prunus salicina is a commercially dominant Asian plum that originated in the Yangtze River basin (Faust and Surányi, 1999). This type of plum is consumed as a fresh fruit and is usually known as the Japanese plum. Prunus simonii, commonly called Simon plum or apricot plum, originated from northern China. Because of its useful traits, such as cold hardiness and firm flesh, it has been used to hybridize with P. salicina (Topp et al., 2012).
Approximately 13-23 species of the American plum group have been reported with the major species being Prunus americana, Prunus hortulana, and Prunus maritima (Rehder, 1954). The American species are adapted to a wide range of environments and have many horticulturally important traits, such as bacterial spot resistance, drought tolerance, and cold hardiness (Topp et al., 2012).
Among these various plum species, P. domestica and P. salicina are major commercial species in the world (Byrne, 2012). Although they belong to the same taxonomic section, they are distinct crops in terms of their uses, adaptation, origin, and domestication. The fruit of P. salicina is primarily used for fresh consumption, whereas that of P. domestica is processed, such as through canning and drying to produce prunes (Topp et al., 2012).
In the Republic of Korea, plums are consumed as fresh fruits and most of the varieties belong to P. salicina. The major cultivars in the Republic of Korea are ‘Ooishiwase’, ‘Formosa’, and ‘Akihime’, which originated from Japan (Chung et al., 2013). ‘Ooishiwase’ is an early-season cultivar that is harvested at around late June, whereas ‘Formosa’ and ‘Akihime’ are harvested in mid July and early September, respectively (Chung et al., 2013). Since 1947, the National Institute of Horticultural and Herbal Science (NIHHS, Rural Development Administration, Republic of Korea) has collected plum varieties as genetic resources for use as parental material in plum breeding programs (NIHHS, 2008). In breeding programs, the selection of superior varieties is extremely important. In order to identify excellent parental materials, comprehensive analyses and evaluations should be carried out in advance. To select superior genetic resources, many studies about germplasm have been done in various crops (Ko et al., 2017; Luitel et al., 2017). For this reason, we evaluated the flower and fruit characteristics of 63 Asian plum varieties and accessions from various countries for three consecutive years. In particular, essential fruit quality factors such as size, shape, skin color, flesh color, soluble solids content (SSC), and acidity were evaluated to select good breeding materials. The objectives of this study were to determine the phenotypic diversity of the selected varieties and to provide helpful guidelines for selecting breeding materials.
Materials and methods
Plant material
Sixty-three varieties and accessions from the plum germplasm collection in NIHHS were evaluated during 2015-17. The species and their origins comprised 12 varieties and accessions from Korea, 23 from Japan, 23 from the USA, and 5 from China (Table 1). The germplasm collection is located in Wanju (35°8′N, 127°0′E), Republic of Korea, which has a temperate climate with an accumulated annual rainfall of approximately 1,300 ㎜ (Korea Meteorological Administration, 2011). Three trees of each variety of this collection were planted in a 6 × 2-m spacing grid. This collection was grafted onto Chinese wild peach seedling rootstocks. Thinning, pruning, irrigation, fertilization, and pest and disease management were performed according to previously published guidelines for plum cultivation (Chung et al., 2013). Dried seed samples of the plum varieties evaluated in this study have been kept as specimens at NIHHS.
Morphological characteristics
Ten morphological characteristics of the flowers and fruits were evaluated according to the Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability for Japanese plum (UPOV, 2011), published by the International Union for the Protection of New Varieties of Plants (UPOV). For the flower characteristics, the petal arrangement, petal shape, petal margin undulation, and stigma position were investigated. For the fruit characteristics, the shape, symmetry, skin color, flesh color, stone adherence to the flesh, and stone shape were investigated. The states and examples of each trait are given in Table 2 and Figs. 1-3.
Table 2. Morphological characteristics used in phenotypic evaluations of plum germplasm (UPOV, 2011) |
|


Fig. 1.
Different shapes in four flower characteristics. A, arrangement of petals; A-1 and A-2, free; A-3 and A-4, touching; A-5 and A-6, overlapping; B, petal shape; B-1 and B-2, elliptic; B-3 and B-4, obovate; B-5 and B-6, circular; B-7 and B-8, oblate; C, undulation of margin in petal; C-1 and C-2, weak; C-3 and C-4, medium; C-5 and C-6, strong; D, stigma position in relation to anthers; D-1 and D-2, below; D-3 and D-4, same level; D-5 and D-6, above.

Fig. 2.
Different shapes and colors of fruit. A, fruit shape; A-1 and A-2, elliptic; A-3 and A-4, circular; A-5 and A-6, oblate; A-7 and A-8, cordate; B, fruit symmetry; B-1 and B-2, symmetric or slightly asymmetric; B-3 and B-4, asymmetric; C, over color of fruit skin; C-1 and C-2, yellow; C-3 and C-4, orange yellow; C-5 and C-6, medium red; C-7 and C-8, dark red; C-9 and C-10, purple; C-11 and C-12, black.

Fig. 3.
Different types of fruit flesh and stones. A, flesh color; A-1 and A-2, whitish; A-3 and A-4, green; A-5 and A-6, yellowish green; A-7 and A-8, yellow; A-9 and A-10, orange; A-11 and A-12, medium red; A-13 and A-14, dark red; B, adherence of stone to fruit flesh; B-1 and B-2, non-adherent; B-3 and B-4, semi-adherent; B-5 and B-6, adherent; C, stone shape in lateral view; C-1 and C-2, narrow elliptic; C-3 and C-4, medium elliptic; C-5 and C-6, circular; C-7 and C-8, broad ovate.
Phenological characteristics
The flowering and ripening times, which are phenological characteristics, were also monitored. The flowering date was recorded as the time when 80% of the flowers are open, and the ripening time was recorded as the time when 20% of the fruits are in the soft stage. These characteristics were measured in Julian days. The fruit development period (FDP) was measured as the interval between the dates of full bloom and ripening.
Fruit characteristics
To evaluate the fruit characteristics, samples of 30 fruits per variety, with 10 fruits from each tree as a replication, were harvested randomly. The fruit weight was measured using a digital scale (MW-2000, Casbee, Yangju, Republic of Korea) in gram (g) units. The fruit height and width were measured with a digital caliper (CD-20CPX, Mitutoyo Corp., Kawasaki, Japan) in millimeter (㎜) units. The SSC was determined with a refractometer (PAL-1, Atago, Tokyo, Japan) in °Brix units. The titratable acidity (TA) was determined by titrating 5 ㎖ of the fruit juice to pH 8.1 with 0.1 N NaOH in an automated titration system (TitroLine Easy™ Module 2, Schott, Mainz, Germany), with the data given in grams per liter (g・L-1) of malic acid, which is the dominant organic acid in plums (Willis et al., 1983). The ripening index (RI) was calculated on the basis of the SSC/TA ratio.
Data analysis
Data are presented as the means of the three years, with three replications each year. Pearson’s correlation coefficients between traits were determined, and all statistical analyses were performed using the R package (version 3.3.1) (R Core Team, 2016).
Results and discussion
The frequency distributions of nine traits are given in Fig. 4. The petal arrangements were grouped as free, touching, and overlapping (Fig. 4A). More than half of the accessions (38) showed the free shape, whereas 18 showed touching and 7 showed overlapping shapes. The petal shapes also varied as elliptic (13), circular (18), oblate (31), and obovate (1) forms (Fig. 4B). With regard to the undulation of the petal margin, most accessions showed a weak (36) or a medium (25) degree of undulation, and only two varieties showed strong undulation (Fig. 4C). These flower shapes can influence the attraction of pollinating insects to the plant.
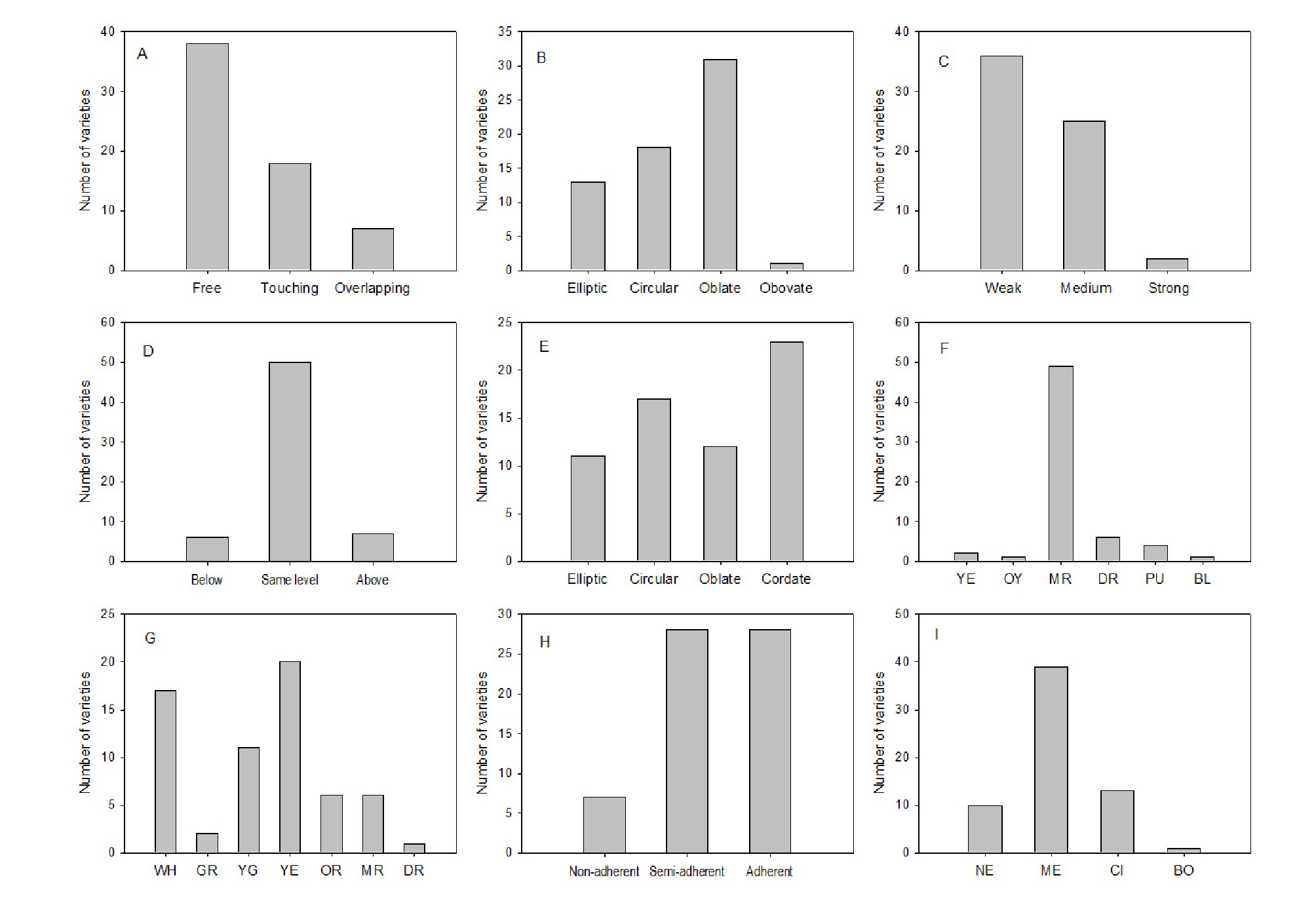
Fig. 4.
Frequency distribution of flower and fruit characteristics. A, Arrangement of petals; B, petal shape; C, undulation of petal margin; D, stigma position in relation to anthers; E, fruit shape; F, over color of skin (YE, yellow; OY, orange yellow; MR, medium red; DR, dark red; PU, purple; BL, black); G, color of fruit flesh (WH, whitish; GR, green; YG, yellowish green; YE, yellow; OR, orange; MR, medium red; DR, dark red); H, adherence of stone to fresh; I, stone shape in lateral view (NE, narrow elliptic; ME, medium elliptic; CI, circular; BO, broad ovate).
Regarding the stigma’s position in relation to the anther’s, most accessions showed them being at the same level (50), whereas six accessions showed a lower stigma level and seven accessions showed a high positioning (Fig. 4D). This flower trait is closely related to pollination and fertilization. Most varieties of P. salicina have a self-incompatible mechanism that prevents self-fertilization (Okie and Hancock, 2008). In terms of the genetics, self-incompatibility is controlled by S-alleles that are expressed in the style and pollen (Kao and Tsukamoto, 2004). When the S-alleles of the pollen are identical to those of the style, pollen tube growth is arrested in the style. Morphologically speaking, the flower could avoid self-pollination through different positioning of the stigmas relative to the anthers in the same flower. Specifically, stigmas that are higher in position to the anthers rarely receive their own pollens (Figs. 1D-5 and 1D-6).
The fruit shapes were grouped as elliptic (11), circular (17), oblate (12), and cordate (23) (Fig. 4E). The elliptic shape is similar to a narrow oval form in which the height is longer than the width (Fig. 2A). The circular shape is round, with similar height and width lengths. The oblate shape is a flat form, with the width being longer than the height. Cordate describes a heart shape, with the stalk attached to the notch. The elliptic, circular, and oblate shapes have the broadest part at the middle of the fruit, whereas the broadest part of the cordate shape is close to the stalk. In this study, the majority of the fruits were cordate and circular in shape. Although the UPOV guidelines include the oblong, obovate, and obcordate shapes, these were not found in this study. This result is in accordance with a report stating that Asian plums are mostly round or heart shaped and never oval or elongated (Hartmann and Neumüller, 2009). In the fresh market in Korea, most plums are also cordate and circular in shape. Although ‘Kelsey’ was not evaluated in this study, the fruit of this cultivar is unique for its long heart shape with an extended apex. This unusual shape among cultivated plums in Korea is one of the reasons why ‘Kelsey’ is cultivated in the country. Therefore, the diversification of fruit shapes can also be a breeding target to increase fruit consumption.
In the case of fruit symmetry, only 16 varieties were symmetric or slightly asymmetric and the other varieties were asymmetric. The symmetric fruit can avoid damage during harvesting and packing (Topp et al., 2012), and is also visually attractive to consumers in the fresh market.
The overall color of the fruit skin was medium red in 49 (78%) of the 63 accessions studied (Fig. 4F). Yellow (2), orange yellow (1), dark red (6), purple (4), and black (1) fruit skin colors were also observed. Although Topp et al. (2012) mentioned that red and black are the most common skin color in Asian plums, red was the major color found in our present study. The skin color is quantitatively inherited, where a yellow skin color is controlled by a single gene that is recessive to black and red (Weinberger and Thompson, 1962). Although the UPOV test guidelines suggested dark blue as a fruit skin color, our collection had no accession with this color. Whereas dark blue is a very common fruit skin color in P. domestica, it is very rare in P. salicina.
Variations were also observed in the fruit flesh color (Fig. 3A), which ranged as whitish (17), green (2), yellowish green (11), yellow (20), orange (6), medium red (6), and dark red (1) (Fig. 4G). Fruit color is determined by the various pigments present in the tissue (Rood, 1957; Stembridge et al., 1972). In this study, six varieties whose pollen parent was ‘Taiyo’ had whitish or yellowish green fruit flesh. In the cases of ‘Wonkyo Ma-03’, ‘Wonkyo Ma-06’, and ‘Wonkyo Ma-15’ (all from seed parent ‘Soldam’ and pollen parent ‘Taiyo’), the flesh color was red. However, the flesh color of ‘Wonkyo Ma-03’, ‘Wonkyo Ma-06’, and ‘Wonkyo Ma-15’ was whitish or yellowish green, which is far from red. It would seem that this bright flesh color came from the whitish flesh of ‘Taiyo’. Owing to the small number of offspring evaluated in this study, further study on the heritability of the flesh color is needed.
Recently, many plum studies on functional chemicals to provide health benefits have been carried out (Gil et al., 2002). Vizzotto et al. (2007) reported that red-fleshed plums generally have a higher anthocyanin and phenolic content than plums with a whitish or a yellow flesh color. Although the fruit skin contains 3-9 times more phenolics than the flesh does, the flesh provides about 70% of the total amount because of its larger fraction (Cevallos-Casals et al., 2006). Sometimes, the extremely high levels of phenolics is associated with astringency (Gil et al., 2002); thus, further studies on the inheritance of key traits are needed.
In the case of stone adherence, only seven varieties showed the non-adherent trait, whereas most of the varieties showed semi-adherent (28) or adherent (28) characteristics (Figs. 3B and 4H). Stone adherence is a quantitative trait, and there are several reports that clingstones seem to be dominant over freestones (Paunović et al., 1968). Although the majority of Asian plums are clingstones, freestone fruits are generally desired for easier consumption. In this study, ‘Fortune’, ‘Kiyou’, and ‘Sunny Queen’ were freestones and these can be used for freestone fruit breeding.
The stone shapes were also observed in lateral view (Fig. 3C). In this study, narrow elliptic (10), medium elliptic (39), circular (13), and broad ovate (1) shapes were evident (Fig. 4I). The shapes of the stones were usually similar to their own fruit shapes (data not shown). For example, ‘Shihou’, whose fruit shape was elliptic, had a narrow elliptic stone.
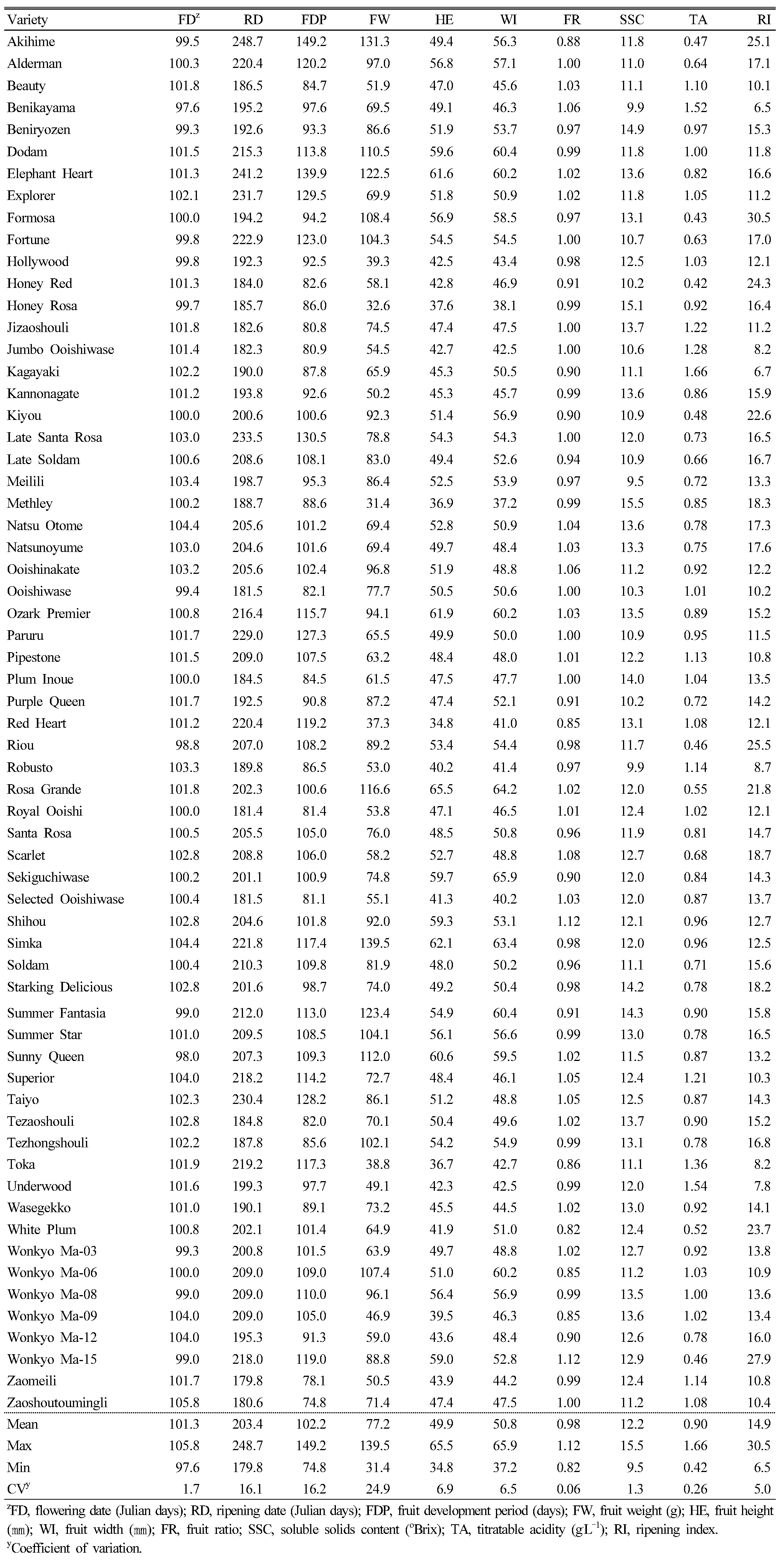
The averages and ranges of 10 phenotypic characteristics with numerical values are shown in Table 3. The average flowering date was April 11th (101 Julian days). The earliest variety to bloom was ‘Benikayama’, which bloomed on April 7th (97 Julian days). The latest cultivar was ‘Zaoshoutoumingli’, which bloomed on April 15th (105 Julian days). The blooming time is an important parameter in many stone fruit breeding programs. To avoid frost damage, late blooming is a breeding target in apricots, which bloom earlier than other stone fruits do (Krška, 2017). Usually, the high-chill genotypes bloom later and they are advantageous in cold regions. On the other hand, in warmer regions, inadequate chilling results in delayed and abnormal flowering, which can reduce productivity. Therefore, low-chill genotypes are needed in warm regions. At the University of Florida, two low-chill plum seedlings whose chilling requirements are only 100 chilling units were selected for planting in the region, which has a warm climate year round (Topp and Sherman, 1990; Sherman et al., 1992). In Korea, plums usually bloom a few days later than apricots do, and the frost damage of plums has not been a serious problem so far.
The information of flowering time is also useful when selecting suitable pollinizers (Kwon et al., 2017). Because most Asian plums are self-incompatible, a lack of pollen transfer can result in low yields. Therefore, self-compatible varieties are also a breeding target, and selecting a suitable pollinizer with synchronized blooming is important in plum production (Beppu et al., 2010).
In this study, the flowering span among the studied varieties was about eight days. In warmer regions, the time span between the early- and late-flowering varieties is longer than that in the cooler regions (LaRue and Norton, 1989). For example, the blooming span among plum varieties was 22 days in Rome, Italy (Hartmann and Neumüller, 2009) where the winter temperature is 8-10°C higher than that in Wanju. If the weather becomes warmer as a result of global warming, the blooming span could be extended. A longer blooming span is an unfavorable condition for pollination because the flowering times of different varieties are various and the chances to synchronize with each other could be reduced. Therefore, the flowering time under various conditions should be monitored carefully.
The average fruit-ripening date was July 22nd (203 Julian days), ranging from June 28th (179 Julian days) to September 5th (248 Julian days, Table 3). The earliest cultivar was ‘Zaomeili’ and the latest was ‘Akihime’. The average FDP was about 102 days, ranging from 74 to 149 days. ‘Zaoshoutoumingli’ showed the shortest FDP, whereas ‘Akihime’ showed the longest one. In the case of ‘Zaoshoutoumingli’, although it was late blooming, the fruit ripened in the early season (late June) with a very short FDP. Therefore, this cultivar seems to be profitable when grown in forced conditions, such as in a greenhouse. The earlier flowering in a greenhouse might produce earlier fruits, which is advantageous for pre-season marketing. Because a better price can be made at the end and especially at the beginning of the harvesting season, expanding the harvest season with various varieties is one of the breeding targets. Bellini et al. (1998) reported the heritability of the ripening date in P. salicina as being 0.44, and the progeny mean was close to the parental mean (Weinberger and Thompson, 1962). Therefore, crossing varieties that have a very short (<80 days) or a very long (>130 days) FDP is one of the strategies to expanding the harvest season. In this study, the FDPs of ‘Zaomeili’, and ‘Zaoshoutoumingli’ were short (<80 days) and those of ‘Akihime’, ‘Elephant Heart’, and ‘Late Santa Rosa’ were long (>130 days). These cultivars are recommended as parent materials for extending the ripening seasons. It is generally regarded that reducing the FDP to less than 75 days is difficult in P. salicina (Topp et al., 2012). The embryos of early-ripening varieties are often underdeveloped or stones are even empty, and hence the number of obtainable seeds and their germination rate are very low. Therefore, the early-ripening variety should be used as a male parent and a mid-season ripening variety as a female parent. Alternatively, the embryo-rescue method can be used.
Owing to their short shelf life, many continuous plum varieties are needed for the fresh market. Ideally, more than one variety for each week is recommended in plum orchards (Hartmann and Neumüller, 2009). In this study, the last variety in season was ‘Akihme’ (248 Julian days) following ‘Elephant Heart’ (241 Julian days), and ‘Late Santa Rosa’ (233 Julian days). Interestingly, there was a nearly empty season of 15 days between ‘Akihime’ and ‘Late Santa Rosa’; there is only one variety, ‘Elephant Heart’, between them. This vacancy is also found in plum production in Korea. Therefore, the late-season varieties that ripen in late August are desperately needed.
The average fruit weight was 77.2 g, ranging from 31.4 to 139.5 g. ‘Akihime’ and ‘Simka’ presented high fruit weight values (>130 g), whereas ‘Hollywood’, ‘Honey Rosa’, ‘Methley’, ‘Red Heart’, and ‘Toka’ showed low ones (<40 g). Additionally, the average fruit height and width were on average 49.9 and 50.8 ㎜, respectively. ‘Elephant Heart’, ‘Ozark Premier’, ‘Rosa Grande’, and ‘Simka’ showed high fruit height values (>60 ㎜), whereas ‘Honey Rosa’, ‘Methley’, ‘Red Heart’, ‘Toka’, and ‘Wonkyo Ma-09’ presented low values (<40 ㎜). On the other hand, ‘Rosa Grande’, ‘Sekiguchiwase’, and ‘Simka’ showed high fruit width values (>60 ㎜), whereas ‘Honey Rosa’, and ‘Methley’ showed low ones (<40 ㎜). These fruit size traits are important in breeding programs. In the case of fruit weight, only a small percentage of progenies are expected to have larger fruits than those of their parents with moderate inheritance (Paunović and Mišić, 1975). Therefore, many progenies are needed to breed larger fruit varieties, and large varieties such as ‘Akihime’ and ‘Simka’ are recommended as parents in crosses.
The width/height ratio was also calculated to investigate the fruit shape, where those with a lower ratio are close to oblate and those with a higher ratio are close to elliptic. In this study, the ratio ranged from 0.82 (‘White Plum’) to 1.12 (‘Shihou’).
Although visual traits such as fruit color, size, and shape influence the consumer’s decision, organoleptic traits such as sweetness, flavor, and texture influence repeated sales. The average SSC was 12.2 °Brix, ranging from 9.5 to 15.5 °Brix. ‘Beniryozen’, ‘Honey Rosa’, ‘Methley’, ‘Starking Delicious’, and ‘Summer Fantasia’ showed high SSC values (>14.0 °Brix), whereas those for ‘Benikayama’ , ‘Meilili’, and ‘Robusto’ were low (<10.0 °Brix). The TA value ranged from 0.42 to 1.66 g・L-1, with an average of 0.90 g・L-1. ‘Benikayama’, ‘Kagayaki’, and ‘Underwood’ had high acidity (>1.5 g・L-1), whereas ‘Akihime’, ‘Formosa’, ‘Honey Red’, ‘Kiyou’, ‘Riou’, and ‘Wonkyo Ma-15’ had low acidity (<0.5 g・L-1<).
Taste attributes, such as sweetness and sourness, are very important in fruit quality. However, these characteristics are affected by the environment and every consumer has different preferences. Crisosto et al. (2004) reported that plums with an SSC of 10.0-11.9 °Brix combined with a low acidity (<0.60%) were acceptable by 56.4% of consumers. Plums with a high SSC (>12.0 °Brix) had approximately 75% consumer acceptance regardless of acidity. Moreover, the preferred taste varies from country to country. Consumers in Asia and South Europe prefer sweet fruits, whereas people in other countries prefer a balanced taste between the sugar and acid contents (Hartmann and Neumüller, 2009).
Correlations between traits
The Pearson’s correlation and each coefficient value between pairs of traits are shown in Fig. 5. The ripening date showed significant and positive correlations with FDP (r = 0.99***), fresh weight (r = 0.42***), fruit height (r = 0.32*), and fruit width (r = 0.30*). According to this result, when fruits are harvested later, they will be larger in size. In general, when fruit are harvested later, they have a larger size and higher SSC. It means that early-ripening cultivars with high quality are not common (Byrne, 2012). In this study, however, there was no relationship between the ripening date and SSC, which is a major fruit quality factor. It seems that several early-season varieties with a high SSC, such as ‘Jizaoshouli’, ‘Plum Inoue’, ‘Royal Ooishi’, ‘Tezaoshouli’, and ‘Zaomeli’ were among this correlation analysis. Moreover, the SSC of the late-season cultivars, such as ‘Akihime’ and ‘Late Santa Rosa’, were not that high (<12.5 °Brix). According to this result, late-season varieties of a high quality are needed.
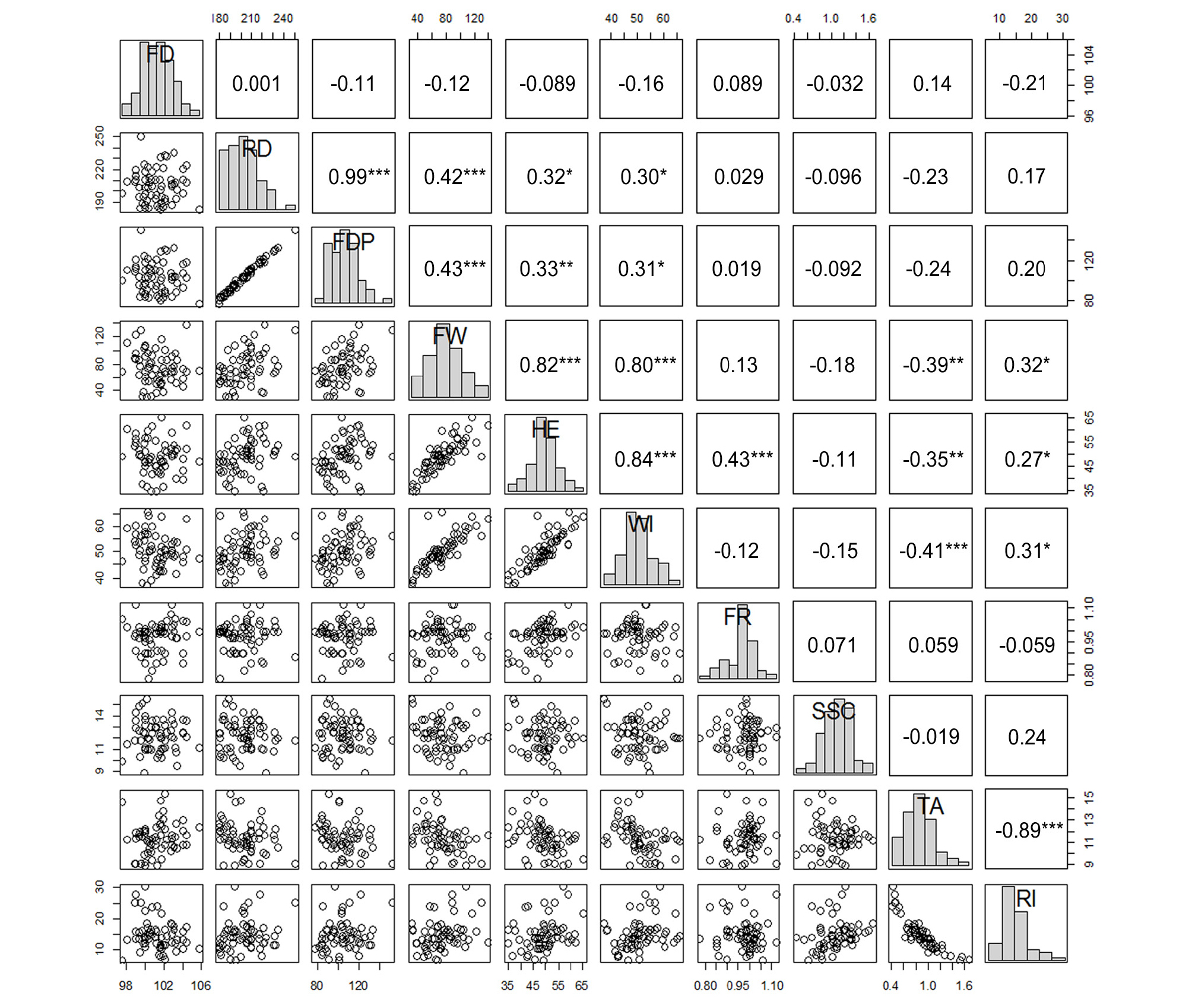
Fig. 5.
Scatter plot matrix and Pearson’s correlation coefficient between pairs of traits studied. FD, flowering date; RD, ripening date; FDP, fruit development period; FW, fruit weight; HE, fruit height; WI, fruit width; FR, fruit ratio; SSC, soluble solids content; TA, titratable acidity; RI, ripening index. *** P ≤ 0.001, ** P ≤ 0.01, * P ≤ 0.05 represent significant values.
Significant positive correlations were also found between the FDP and fruit weight (r = 0.43***), fruit height (r = 0.33**), and fruit width (r = 0.31*). Moreover, the fruit size factors of weight, height, and width showed positive correlations with one another. The TA value showed negative correlations with the fruit weight (r = -0.39**), fruit height (r = -0.35**), and fruit width (r = -0.41***). This indicates that larger fruits have a lower TA value. When the breeding target is a large fruit with low acidity, ‘Akihime’ and ‘Formosa’ (with low TA values and large fruit sizes) are recommended as breeding materials. The SSC had no correlation with any other characteristics, whereas the RI value showed negative correlation with TA (r = -0.89***) and fruit size traits. The RI value, which is the sugar/acid ratio, is commonly used as a quality index (Robertson et al., 1989; Bassi and Selli, 1990) and would be a useful trait for estimating fruit taste.
In this study, 63 plum varieties collected from various countries were evaluated to determine their phenotypic diversity and relationships. To select superior breeding materials, various phenotypic traits were thoroughly evaluated, especially those of fruit quality factors, such as size, shape, skin color, flesh color, SSC, and acidity. Five plum cultivars, ‘Purple Queen’ (2001), ‘Honey Red’ (2002), ‘Summer Fantasia’ (2012), ‘Sunny Queen’ (2013), and ‘Summer Star’ (2016), were released from the NIHHS. The main target in plum breeding programs is a fruit of high quality, with a high SSC and large fruit size, and attempts are ongoing to find superior breeding materials to achieve these traits. Diverse varieties provide breeders with a wider range of breeding materials. At this moment, about 79% of the plum germplasms in the NIHHS belong to P. salicina. To expand diversification, the collection of other plum species, such as Prunus cocomila and Prunus munsoniana, is strongly recommended. P. salicina also has advantages in diversification because this species is compatible with other species, such as P. cerasifera, P. simonii, P. americana, P. hortulana, and P. munsoniana (Hedrick, 1911; Howard, 1945). Byrne (1990) also mentioned that the diploid Asian plums have greater diversity than peaches, which are self-pollinating single species. Through this study, information of plum varieties has been provided for aiding the future selection of superior parents. Furthermore, the correlations observed among the fruit quality parameters should simplify the evaluation procedures.
