

서 언
재료 및 방법
재료
신서란 추출물 및 분획물 제조
세포 배양
세포 독성 측정
Nitric oxide(NO) 생성 억제 활성 측정
Prostaglandin E2(PGE2) 생성 억제 활성 측정
염증성 사이토카인(TNF-α, IL-6, IL-β) 생성 억제 활성 측정
iNOS 및 COX-2 발현 억제 효능 평가(Western blot analysis)
Chemokine(MDC, TARC) 생성 억제 활성 측정
통계분석
결과 및 고찰
에탄올 추출물과 용매 분획물의 수율
세포 독성에 미치는 영향
NO 및 Prostaglandin E2(PGE2) 생성 억제 효과
iNOS 및 COX-2 단백질 발현에 미치는 영향
Pro-inflammatory cytokine(TNF-α, IL-6, IL-1β) 생성 억제 활성
Chemokine 생성 변화에 미치는 영향
적 요
서 언
염증은 물리화학적 자극이나 세균 감염에 대한 생체조직의 방어 반응의 하나이며, 조직이나 장기의 손상을 회복시키는 기전으로서 매우 중요한 역할을 담당하고 있다(Zamora et al., 2000). 그러나 염증반응이 과도해질 경우 동맥경화, 류마티스 관절염, 천식, 기관지염, 암 등을 유발하는 원인으로 작용하기 때문에 염증반응을 조절하는 항염증제 개발은 매우 중요시되고 있다(Kaplanski et al., 2003, Ko et al., 2015). 염증반응이 일어나면 대식세포와 같은 염증세포들은 nitric oxide (NO), prostagladin E2 (PGE2), tumor necrosis factor-α (TNF-α) 및 interleukin-6 (IL-6) 등과 같은 염증매개 물질을 분비 한다(Lazarov et al., 2000). 또한, inducible nitric oxide synthase (iNOS)와 cyclooxygenase-2 (COX-2)의 발현을 유도함으로서 NO와 PGE2 등이 생성된다(Albina and Reichner, 1999). 염증반응이 지표물질인 NO는 nitric oxide synthase (NOS)에 의하여 생성이 조절되는데(Moeslinger et al., 2006), iNOS는 lipopolysaccharide (LPS) 등의 자극을 통해 발현되어 많은 양의 NO를 생성한다(Guzik et al., 2003). 또한, IL-1, TNF-α, interferon-γ (IFN-γ)와 같은 전염증성 cytokine의 형성은 arachidonic acid가 cyclooxygenase (COX)의 작용을 거쳐 prostaglandin 등으로 바뀌는 과정에 관여함으로써 염증 매개에 큰 역할을 하는 것으로 알려져 있다(Guo et al., 2012).
한편, 아토피 피부염(atopic dermatitis)은 유전, 환경 및 식품 등 다양한 요인들에 의해 복합적으로 발생하는 염증성 피부질환으로서(Guttman-Yassky et al., 2011; Shaw et al., 2011), 알레르기 유발물질이 피부에 침입 할 경우 제2형 보조 T 세포(T-helper2, Th2)에 의해 IL-4, IL-13 및 IL-1β와 같은 cytokine이 분비되고, 표피에서는 thymus & activation-regulated chemokine (TARC/CCL17)과 macrophage-derived chemokine (MDC/CCL22)에 의해 Th2 세포가 피부 병변으로 유도되어 발병하는 것으로 알려져 있다(Gandhi et al., 2017).
최근 들어 건강과 뷰티 관련 바이오산업이 급성장하면서 천연물 혹은 생약의 유효성에 대한 연구가 활기를 띠고 있는데, 특히 염증성 질환 예방 및 치료를 위한 소재 발굴이 활발하게 이루어지고 있다. 어성초(Houttuynia cordate), 오가피Acanthopanacis cortex), 황기(Astragalus membranaceus), 천문동(Asparagus cochinchinensis) 등과 같은 전통 한약재뿐만 아니라 까마귀쪽나무(Litsea japonica) 와 같은 특용작물들은 염증세포의 활성을 조절하고, 관련 사이토카인 발현을 억제하는 항염증 및 항산화 효과가 뛰어난 것으로 알려져 있다(Ham et al. 2017; Kim and park, 2006; Kim et al., 2013; Lee et al., 2008; Woo et al., 2018; Yoon et al., 2015).
신서란(phormium tenax)은 마섬유의 일종으로 뉴질랜드가 원산지이며, 높이는 1~2 m로서 옆으로 뻗은 뿌리줄기에서 긴 칼 모양의 잎을 갖고 있는데, 잎은 끝이 뽀족하고 길이가 1~3 m, 폭이 5~10 ㎝ 이상이며 섬유질이 잘 발달한 식물로서 제주지역에서 오래전부터 자생하고 있다(Kim et al., 2002). 뉴질랜드 마오리어로 하라케케(Harakeke) 불리우는 신서란은 오랫동안 마오리 족에 의해 의복을 만들거나 전통가옥의 지붕, 돗자리, 어망 등을 만드는데 사용되었으며, 여러 증상의 효과적인 약용식물로 널리 사용되어 왔다. 특히, 신서란의 줄기와 잎 사이에서는 플렛스검(Flaxgum)이라 불리우는 알로에 베라와 같은 투명 젤리 모양의 수액이 생산되는데, 이 harakeke flaxgum은 천연 살균제로서 피부종기, 화상, 습진 상처 등에 효과적으로 사용되었다(Riley, 1994). 최근에는 비누, 핸드크림, 샴푸 등과 같은 향장품에 적용하기 위한 특허 등록도 이루어지고 있다(Joung et al, 2018; Kang et al., 2017).
본 연구에서는 하라케케로 알려진 신서란이 항염증 또는 항아토피의 특성을 갖는 특용자원으로서 활용 가치 여부를 평가함으로서 화장품 및 의약품산업에 기능성 소재로써 적용 가능성을 확인하고자 하였다. 이를 위해 신서란 잎을 대상으로 추출물과 극성에 따라 순차적으로 용매 분획한 분획물을 제작하였고, 이렇게 제작된 시료를 LPS로 자극된 RAW 264.7 세포에 처리한 후 염증성 매개 인자인 NO와 PGE2의 생성 억제 효과 및 이를 합성하는 iNOS와 COX-2단백질 발현 억제 효능을 확인하였고, 염증성 cytokine(TNF-α, IL-6)의 생성 억제 효과를 조사하였다. 아토피 질환에 대한 활용 가능성 여부는 각질형성세포인 HaCaT cell에서 아토피 유발인자로 알려진 chemokine (MDC, TARC) 생성 억제효과를 조사하였다.
재료 및 방법
재료
제주특별자치도 서귀포시 남원읍 신례리 일대에서 2015년 8월에 채집한 신서란 잎을 열풍건조(40℃, 72 h) 시킨 후, 마쇄기로 분쇄하여 미세 분말화 한 후 4℃에 보관하면서 추출용 시료로 사용하였다. 채집 신서란은 백합과에 속하는 Phormium tenax로서 (재)제주테크노파크 생물종다양성연구소 고영순 선임연구원님의 식물학적 동정을 거쳤으며, 실험에 사용한 시료의 확증표본(표본번호 P15-061)은 (재)제주테크노파크 생물종다양성연구소에 보관하고 있다.
신서란 추출물 및 분획물 제조
신서란 잎 분말(400 g) 시료를 70% 에탄올(Daejung, Korea)을 이용하여 상온에서 24시간 동안 3회 침출한 후, 감압농축기(laborota 4000, heidoph, Germany)로 감압 농축하여 에탄올 추출물을 얻었다. 에탄올 추출물 5 g을 증류수 500 mL에 현탁 시킨 후, n-hexane (500 mL × 2, Daejung, Korea), methylene chloride (500 mL × 2, Daejung, Korea), ethyl acetate (500 mL × 2, Daejung, Korea), n-butanol (500 mL × 2, Daejung, Korea)로 순차적으로 분획 및 농축하여 각각의 분획물을 확보한 후 동결건조기(7960044, Labconco, USA)로 동결 건조하여 n-hexane, methylene chloride, ethyl acetate, n-butanol 및 수용액 분획물을 얻었다. 제조된 추출물 및 분획물은 –20℃에서 보관하면서 사용하였다.
세포 배양
Murine macrophage cell line인 RAW 264.7 세포를 한국 세포주 은행(Korean Cell Line Bank)으로부터 구입하여 10% fetal bovine serum (FBS)와 100 units/mL penicillin-streptomycine이 함유된 Dulbecco’s Modified Eagle Medium (DMEM, GIBCO, USA) 배지를 사용하여 37℃, 5% CO2 incubator (HERACell 240, ThermoFisher Scientific, USA)에서 3일 간격으로 배양을 하였다. 항 아토피 실험에 사용한 human keratinocyte cell line인 HaCaT cell 역시 10% FBS과 100 unit/mL penicillin-streptomycine을 첨가한 DMEM 배지에 넣고 37℃, 5% CO2 항온기에서 배양하였으며 3일에 한번 씩 계대배양을 하였다.
세포 독성 측정
신서란 조추출물 및 용매 분획물이 세포 성장에 미치는 영향을 측정하기 위하여 lactate dehydrogenase (LDH) assay를 시행하였다. RAW 264.7 세포(1.8 × 105 cells/mL)를 10% FBS가 첨가된 DMEM배지를 이용하여 24 well plate에 분주한 후 24시간 동안 배양한 후 시료와 LPS (1 ㎍/mL)를 동시에 처리하고 24시간 동안 배양하였다. 이후 배양 배지를 3,000 rpm에서 5분간 원심 분리하여 상등액을 취하였다. LDH assay는 non-radioactive cytotoxicity assay kit (CytoTox96, Promega, WI, USA)를 이용하여 측정하였다. 96 well plate에 원심분리하여 얻은 배양 배지 50 μL와 reconstituted substrate mix 50 μL를 넣고, 실온에서 30분 반응시킨 후 50 μL stop solution을 넣고 microplate reader (PowerWave XS2, Bio-TEK instrument Inc., Vermont, WI, USA)를 사용하여 490 ㎚에서 흡광도를 측정하였다.
Nitric oxide(NO) 생성 억제 활성 측정
RAW 264.7 cell을 10% FBS가 첨가된 DMEM배지를 이용하여 1.8 x 105 cells/mL로 조절 한 후 24 well plate에 넣고 18시간 배양하였다. 그 다음 시료와 LPS (1 ㎍/mL)를 동시에 처리하여 24시간 배양하였다. 생성된 NO의 양은 Griess 시약[1% (w/v) sulfanilamide, 0.1%(w/v) naphthylethylendiamine in 2.5% (v/v) phosphoric acid]을 이용하여 세포배양액 중에 존재하는 NO2-의 형태로 측정하였다. 세포배양 상등액 100 μL와 Griess 시약 100 μL를 혼합하여 96 well plates에서 10분 동안 반응시킨 후 540 ㎚에서 흡광도를 측정하였고, 생성된 NO의 양은 sodium nitrite (NaNO2)를 standard로 사용하여 정량하였다.
Prostaglandin E2(PGE2) 생성 억제 활성 측정
RAW 264.7 세포를 DMEM 배지를 이용하여 1.8 × 10⁵ cell/mL로 조절한 후 24 well plate에 접종하고, 5% CO2 항온기에서 18시간 배양하였다. 이후 배지를 제거하고 10배 농도(1 ㎎/mL)로 조제된 시료 50 μL와 LPS (1 ㎍/mL)를 함유한 450 μL의 배지를 동시에 처리하여 24시간 배양하였다. 24시간 후 배양 배지를 원심분리(1,2000 rpm, 3min) 하여 얻어진 상층액의 PGE₂함량을 측정하였다. PGE₂는 mouse enzyme-linked immunosorbent assay (ELISA) kit (R&D System Inc., Minneapoils, MN, USA)를 이용하여 정량하였으며 standard에 대한 표준곡선의 r²값은 0.99 이상이었다.
염증성 사이토카인(TNF-α, IL-6, IL-β) 생성 억제 활성 측정
RAW 264.7 세포를 DMEM 배지를 이용하여 1.8✕10⁵ cell/mL로 조절한 후 24 well plate에 접종하고, 5% CO2 항온기에서 18시간 배양하였다. 이후 배지를 제거하고 10배 농도(1 ㎎/mL)로 조제된 시료 50 μL와 LPS (1 ㎍/mL)를 함유한 450 μL의 배지를 동시에 처리하여 24시간 배양하였다. 24시간 후 배양 배지를 원심분리(1,2000 rpm, 3min) 하여 얻어진 상층액의 pro-inflammatory cytokine 생성 함량을 측정하였다. 전염증성 cytokine은 mouse enzyme-linked immunosorbent assay (ELISA) kit Invitrogen, USA)를 이용하여 정량하였으며 standard에 대한 표준곡선의 r²값은 0.99 이상이었다.
iNOS 및 COX-2 발현 억제 효능 평가(Western blot analysis)
RAW 264.7세포(1.0 × 106 cell/mL)를 18시간 전 배양한 후, 시료와 LPS (1 ㎍/mL)를 동시 처리하여 24시간 배양하였다. 세포를 PBS (phosphate buffered saline)로 2회 세척한 후 200 μL의 lysis buffer [50 mM Tris-HCl (pH 7.5), 150 mM Nacl, 1% Nonidet P-40, 2 mM EDTA, 1 mM EGTA, 1 mM NaVO3, 10 mM NaF, 1 mM dithiothreitol, 1 mM phenyl methyl sulfonyl fluoride, 25 ㎍/mL aprotinin, 25 ㎍/mL leupeptin]를 첨가하여 4℃에서 1시간 동안 lysis시킨 후 15,000 rpm에서 15분간 원심 분리하여 세포막 성분 등을 제거하였다. 단백질 농도는 BSA (bovine serum albumin)를 표준화하여 Bio-Rad Protein Assay Kit (BIO-RAD, USA)을 사용하여 정량하였다. 분리된 단백질 20~30 ㎍을 10~12% mini gel SDS-PAGE (Poly Acrylamide Gel Electrophoresis)로 변성 분리하여, 이를 PVDF (polyvinylidene difluoride) membrane (BIO-RAD, Richmond, USA)에 200 ㎃로 2시간 동안 transfer하였다. 그리고 membrane의 bloking 처리는 5% skim milk가 함유된 TTBS (TBS + 0.1% Tween 20) 용액에서 2시간 동안 실시하였다. iNOS의 발현 양을 확인하기 위한 항체로는 anti-mouse iNOS (1:1000) (Santa- Cruz Biotechnology, USA)을, COX-2의 발현 양을 확인하기 위한 항체로는 anti-mouse COX-2 (BD Biosciences Pharmingen, San Jose, CA, USA)를 TTBS용액에 희석하여 상온에서 2시간 반응시킨 후 TTBS로 4회 세정하였다. 반응이 완료된 membrane을 ECL 기질(Intron Biotechnology, Inc, Korea)과 1분 간 반응 후 X-ray 필름에 감광 하였다
Chemokine(MDC, TARC) 생성 억제 활성 측정
HaCaT cell을 DMEM 배지를 이용하여 5 × 105 cell/mL로 조절 후 24 well plate에 접종하고 5% CO2 항온기에서 18시간 배양하였다. 이후 배지를 제거하고 10배 농도로 조제된 시험물질 50 μL를 hIFN-γ(최종농도 10 ng/㎖)를 포함하는 새로운 배지(FBS(-)) 450 μL에 가하여 전 배양과 동일 조건에서 배양하였다. 24시간 후 배양 배지를 원심분리(12,000rpm, 3min)하여 얻어진 상층액의 MDC 및 TARC 함량을 각각 human MDC immunoassay, human TARC immunoassay kit (R&D System Inc. Minneapoils, MN, USA)를 이용하여 측정하였다.
통계분석
모든 실험은 3반복 이상 실시하였고, 실험결과는 각 항목에 따라 평균치 ± 표준편차(SD)를 구하여 신뢰수준 95% (P<0.05)에서 통계적 유의차를 평가하였다.
결과 및 고찰
에탄올 추출물과 용매 분획물의 수율
신서란 건조 분말시료 400 g을 70% EtOH로 추출한 후 여과하여 얻어진 추출액을 감압 농축하여 조추출물을 확보하였다. 에탄올 추출물(5 g)을 500 mL증류수에 현탁 시킨 후 n-hexane, methyl chloride, ethyl acetate, n-butanol 등으로 순차적으로 분획하여 n-hexane 층에서 112 ㎎ (2.24%), methyl chloride 층에서400 ㎎(8.0%), ethyl acetate 층에서 172 ㎎ (3.44%), n-butanol 층에서 750 ㎎ (15.0%) 및 전사인 물 층에서 3,566 ㎎ (71.32%)의 분획물을 얻었다. 70% 에탄올 추출물에 대한 용매 분획물 중 n-hexane 분획물 수율이 2.24% 로 가장 낮았고, 물 분획물이 71.32%로 가장 높은 수율을 보였다.
세포 독성에 미치는 영향
모든 세포의 세포질 안에 존재하는 효소 LDH는 pyruvic acid와 lactic acid 간의 가역적 반응에 관여하는데, LDH를 내포한 조직이 파괴될 때 혈액 중으로 흘러나와 혈중 LDH가 상승한다. 이 원리를 활용하여 신서란 RAW 264.7 세포에 대상 물질과 LPS (1 ㎍/mL)를 동시에 처리하여 24시간 배양한 후 LDH 분석을 통해서 세포 생존율을 확인한 결과, n-butanol 분획물인 경우 100 ㎍/㎖ 이상의 농도에서 다소 세포 독성이 관찰되었으나 다른 용매 분획물에서는 세포독성이 나타나지 않았고 100 ㎍/mL의 농도 이하에서는 신서란 잎 조추출물 및 용매 분획물에서 세포독성이 나타나지 않았다(Fig. 1).
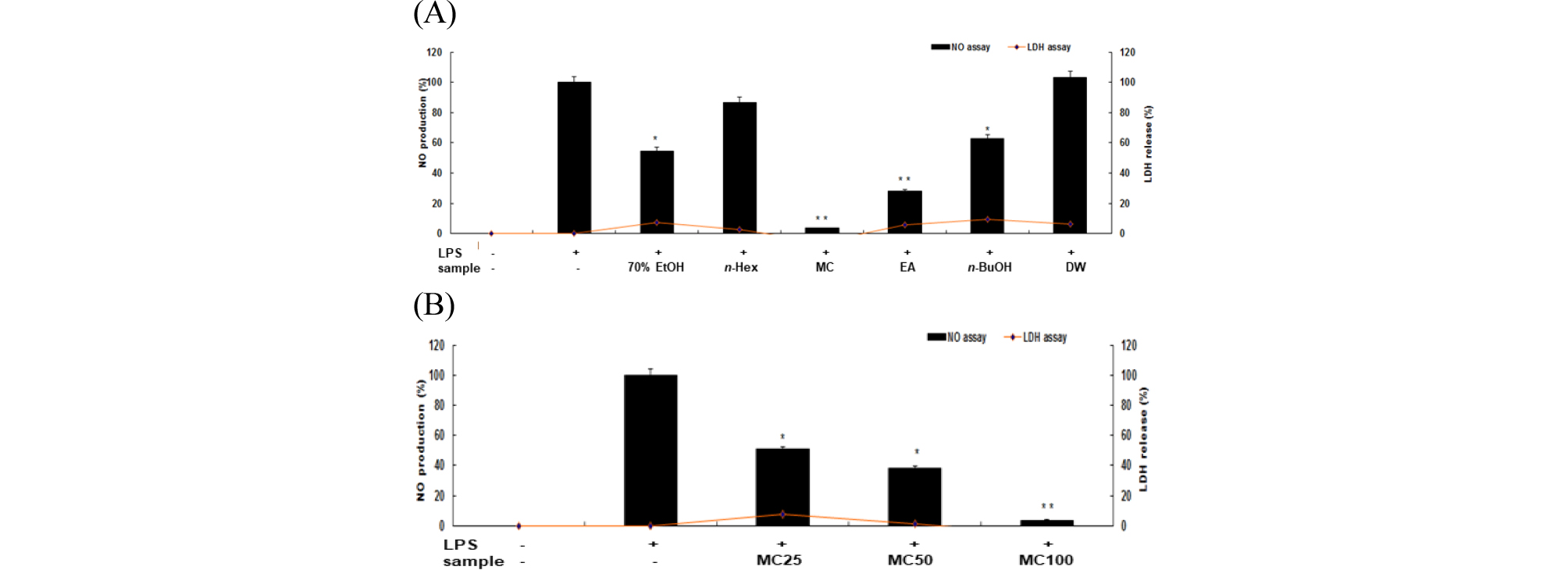
Fig. 1.
Inhibitory effect of crude extract and solvent fractions from P. tenax leaf on NO production and cytotoxicity in LPS-stimulated RAW 264.7 cells. (A): The production of nitric oxide(NO) was assayed in the culture medium of cells treated with LPS (1 ㎍/mL) for 24h in the presence of crude extract and solvent fractions (100 ㎍/mL) of P. tenax leaf. Cytotoxicity was determined using the LDH method. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01. (B) : Cells were treated by LPS (1 ㎍/mL) for 24h in the presence of CH2Cl2 (MC) fractions (25, 50 and 100 ㎍/mL).
NO 및 Prostaglandin E2(PGE2) 생성 억제 효과
대식세포에서는 LPS와 같은 외부자극에 의하여 염증반응이 일어날 때 NO, PGE2, pro-inflammatory cytokines와 같은 물질들이 생성됨으로(Lazarov et al., 2000), 본 연구에서는 신서란 잎 조추출물과 용매 분획물의 NO와 PGE2의 생성에 미치는 영향을 조사하였다. LPS로 자극된 RAW 264.7 세포에 생성되는 NO 양은 Griess 시약을 이용하여 세포 배양액 중에 존재하는 NO2- 의 형태로 측정한 결과, 신서란 잎 조추출물과 용매 분획물에서 NO 생성 억제 효과를 보였는데, 그 중 methylene chloride 분획물과 ethyl acetate 분획물에서 대조군인 LPS 단독처리군에 비해 NO 생성 억제 효과가 크게 나타났다(Fig. 1A). 또한, 가장 활성이 높은 methyl chloride 분획물의 경우 농도 의존적으로 NO 생성 억제 활성을 보였다(Fig. 1B). 한편, prostaglandin (PG)은 arachidonic acid 로부터 유래된 불포화지방산에 속하는 호르몬으로서, 이들 생성된 PG 중 PGE2는 염증반응에 깊게 관여한다. 이에 신서란 잎 조추출물과 용매 분획물에 대한 PGE2 감소 효능을 조사하였다. LPS 무처리군의 PGE2 생성을 0%, LPS 단독 처리군의 PGE2 생성을 100%로 하여 추출물과 분획물 처리군을 비교한 결과, methylene chloride 분획물에서 PGE2 생성억제 효능이 95% 이상, ethyl acetate 분획물에서는 80% 수준임을 확인하였다(Fig. 2A). 이들 두 분획물을 농도별(25, 50, 100 ㎍/mL)로 처리하여 PGE2 생성 억제 활성을 조사한 바, 농도 의존적으로 PGE2 생성이 억제되었다(Fig. 2B).
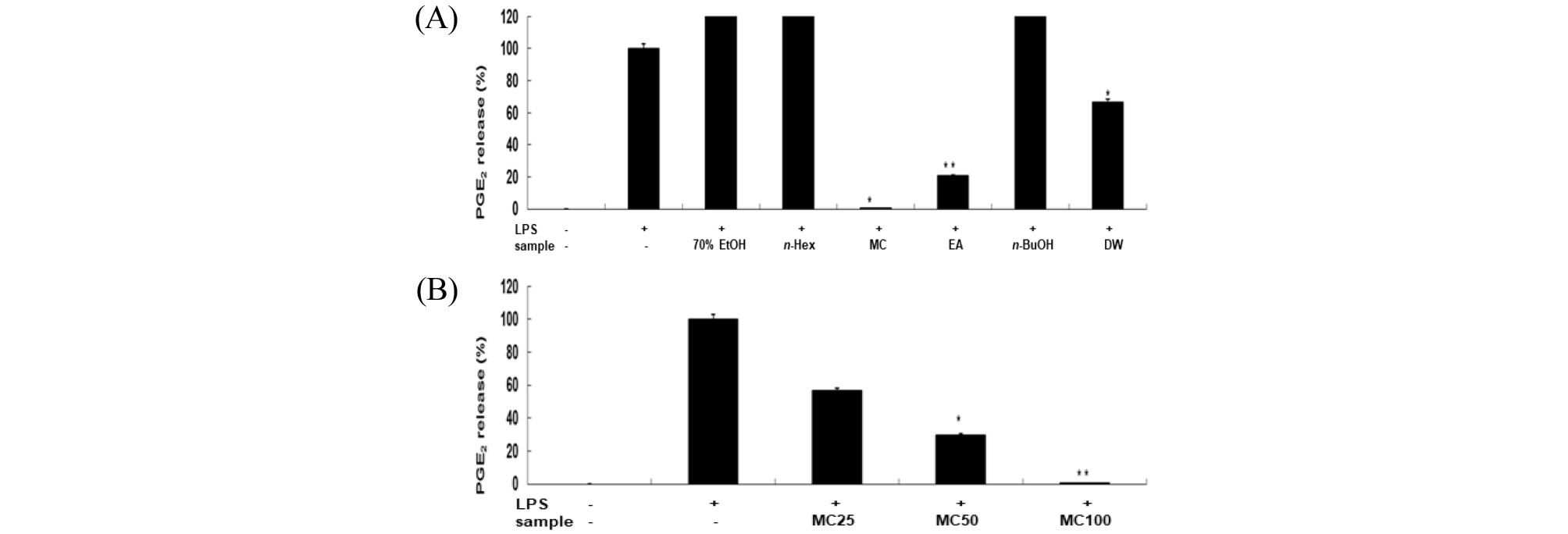
Fig. 2.
Inhibitory effect of crude extract and solvent fractions of P. tenax leaf on PGE2 production in LPS-stimulated RAW 264.7 cells. (A): Cells(1.8×105 cells/mL) were stimulated by LPS(1 ㎍/mL) for 24h in the presence of crude extract and solvent fractions(100 ㎍/mL) of P, tenax leaf. Supernatants were collected, and the PGE2 concentration in the supernatants was determined by ELISA. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01. (B) : Cells were treated by LPS(1 ㎍/mL) for 24h in the presence of CH2Cl2 (MC) fractions(25, 50 and 100 ㎍/mL).
iNOS 및 COX-2 단백질 발현에 미치는 영향
염증반응이 일어나면 대식세포와 같은 염증세포들은 NO, PGE2, TNF-α 및 IL-6 등과 같은 염증매개 물질을 분비 하며(Lazarov et al., 2000), 또한 iNOS와 COX-2의 발현을 유도함으로서 NO와 PGE2 가 생성되게 된다(Albina and Reichner, 1999). Fig. 1과 2에 나타난바와 같이 신서란 추출물 및 분획물에 의한 NO, PGE₂생성 억제 활성이 염증 유발 단백질로 알려진 iNOS와 COX-2 단백질의 발현 억제에 의한 것인지를 알아보기 위하여 단백질 수준에서의 발현을 western blot 방법으로 조사하였다. RAW 264.7 cell에 LPS (1 ㎍/mL) 신서란 잎 조추출물 및 분획물(100 ㎍/mL)를 24시간 처리한 후 iNOS, COX-2의 발현 억제 활성을 확인하였다. 그 결과 LPS 단독 처리군 에서는 iNOS, COX-2의 발현이 현저히 증가하였고 methylene chloride 분획물과 ethyl acetate 분획물에서 iNOS, COX-2의 발현을 억제함을 알 수 있었다(Fig. 3A), 또한, 이들 분획물은 농도(25, 50, 100 ㎍/mL) 의존적으로 iNOS와 COX-2의 발현을 억제하는 효과를 보였다(Fig. 3B). Fig. 3의 결과는 NO 생성 억제가 iNOS 발현 억제를 통한 것으로 여겨진다.
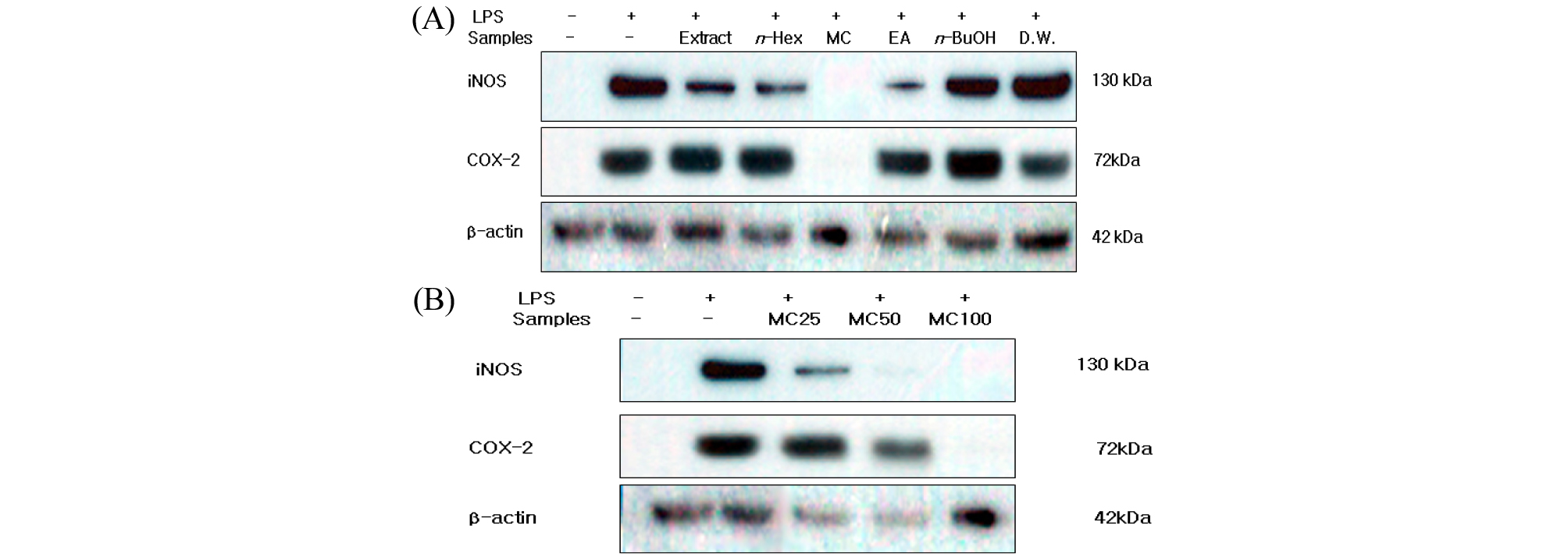
Fig. 3.
Inhibitory effects of crude extract and solvent fractions of P. tenax leaf on the protein level of iNOS and COX-2 in LPS-stimulated RAW 264.7 cells. (A): RAW 264.7 cells(1 × 106 cells/mL) were pre-incubated for 18 hr, and the cells were treated with LPS(1 ㎍/mL) in the presence of crude extract and solvent fractions(100 ㎍/mL) of P, tenax leaf for 24 hr. iNOS and COX-2 protein levels were determined using immunoblotting method. (B): Cells were treated by LPS(1 ㎍/mL) for 24h in the presence of CH2Cl2 (MC) fractions(25, 50 and 100 ㎍/mL).
Pro-inflammatory cytokine(TNF-α, IL-6, IL-1β) 생성 억제 활성
Cytokine은 면역세포가 분비하는 단백질로서 면역세포의 활성, 증식 및 분화를 조절하여 염증반응을 매개하는 인자이다(Guo et al., 2012). Pro-inflammatory cytokine으로 알려진 TNF-α, IL-1β 및 IL-6은 염증반응을 조절하는 물질로 잘 알려져 있다(Delgado et al., 2003; Van Snick. 1990). RAW 264.7세포에서 신서란 잎 조추출물 및 분획물의 염증성 cytokine인 TNF-α, IL-1β 및 IL-6 의 발현에 미치는 영향을 ELISA kit를 이용하여 조사하였다. RAW 264.7 세포에 신서란 추출물 또는 용매 분획물과 LPS (1 ㎍/mL)를 동시 처리하여 24시간 배양 한 후 TNF-α의 생성 억제 활성을 측정한 결과, methylene chloride와 ethyl acetate 분획물에서 각각 70%, 50% 수준으로 억제되고 있음을 보였다(Fig. 4A). IL-6의 생성 억제도 두 분획물에서 각각 85%, 50% 수준으로 억제되는 것으로 나타났고, 농도 의존적으로 억제효능을 보였다(Fig. 5). 또한, IL-1β의 발현에 미치는 영향을 조사한 결과에서도 methylene chloride 와 ethyl acetate 분획물은 각각 99%, 90% 수준으로 뚜렷하게 IL-1β 생성을 억제하는 것으로 관찰되었다(Fig. 6).
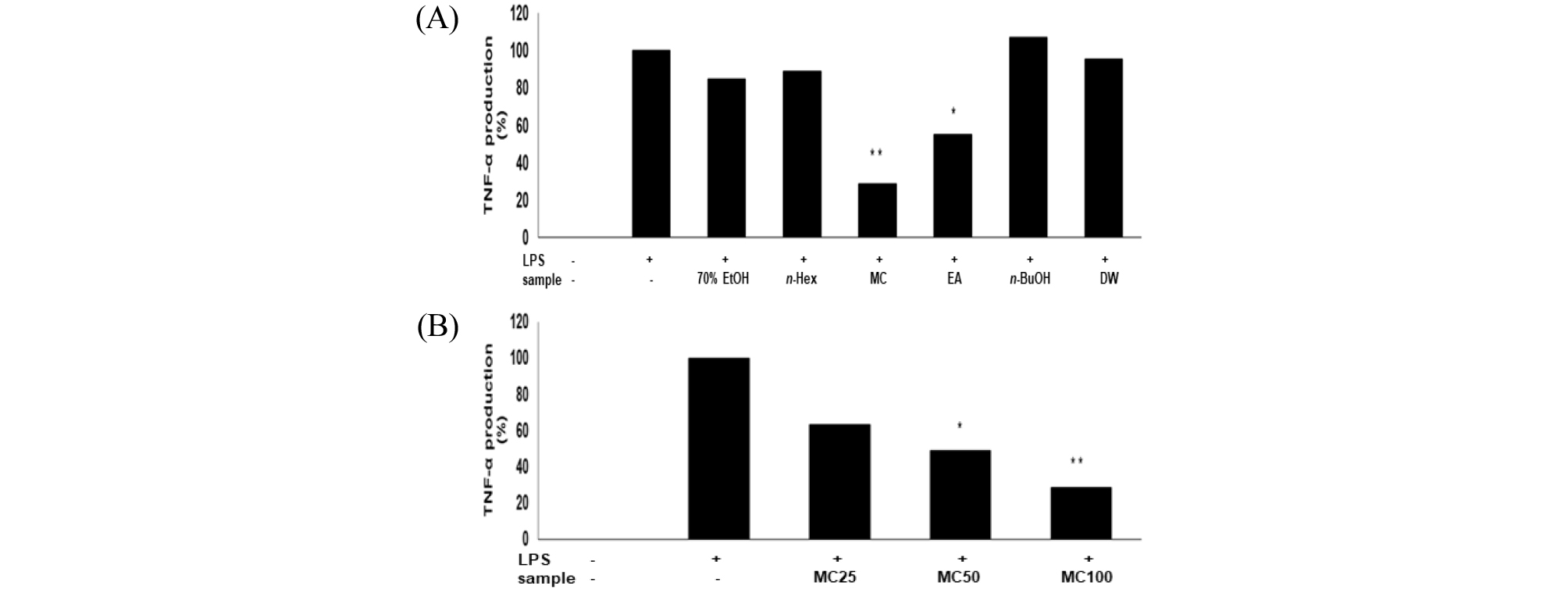
Fig. 4.
Inhibitory effects of crude extract and solvent fractions of P. tenax leaf on TNF-α production in LPS-stimulated RAW 264.7 cells. (A): Cells(1.8 × 105 cells/mL) were treated by LPS (1 ㎍/mL) for 24 h in the presence of crude extract and solvent fractions(100 ㎍/mL) of P. tenax leaf. Supernatants were collected, and the TNF-α concentration in the supernatants was determined by ELISA. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01 (B): Cells(1.8 × 105 cells/mL) were treated by LPS(1 ㎍/mL) for 24 h in the presence of CH2Cl2 (MC) fractions(25, 50 and 100 ㎍/mL).
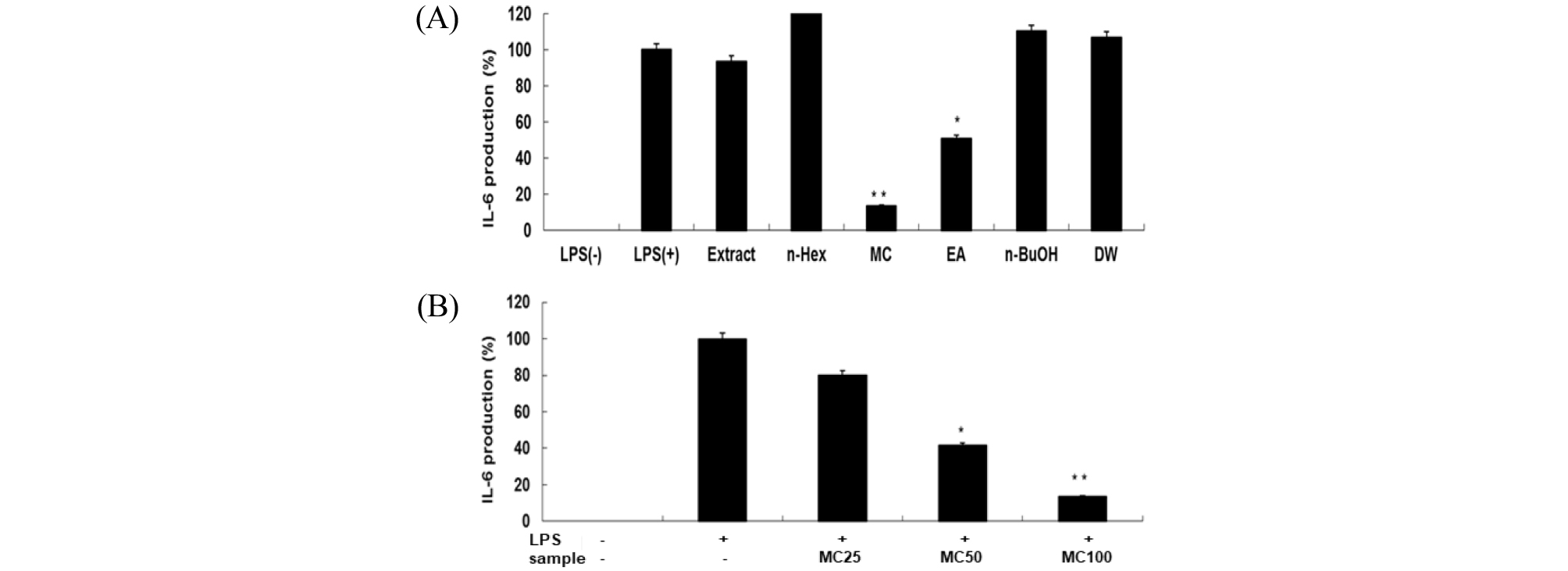
Fig. 5.
Inhibitory effects of crude extract and solvent fractions of P. tenax leaf on IL-6 production in LPS-stimulated RAW 264.7 cells. (A): Cells(1.8 × 105 cells/mL) were treated by LPS(1 ㎍/mL) for 24 h in the presence of crude extract and solvent fractions(100 ㎍/mL) of P. tenax leaf. Supernatants were collected, and the IL-6α concentration in the supernatants was determined by ELISA. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01 (B): Cells were treated by LPS(1 ㎍/mL) for 24 h in the presence of CH2Cl2 (MC) fractions (25, 50 and 100 ㎍/mL).
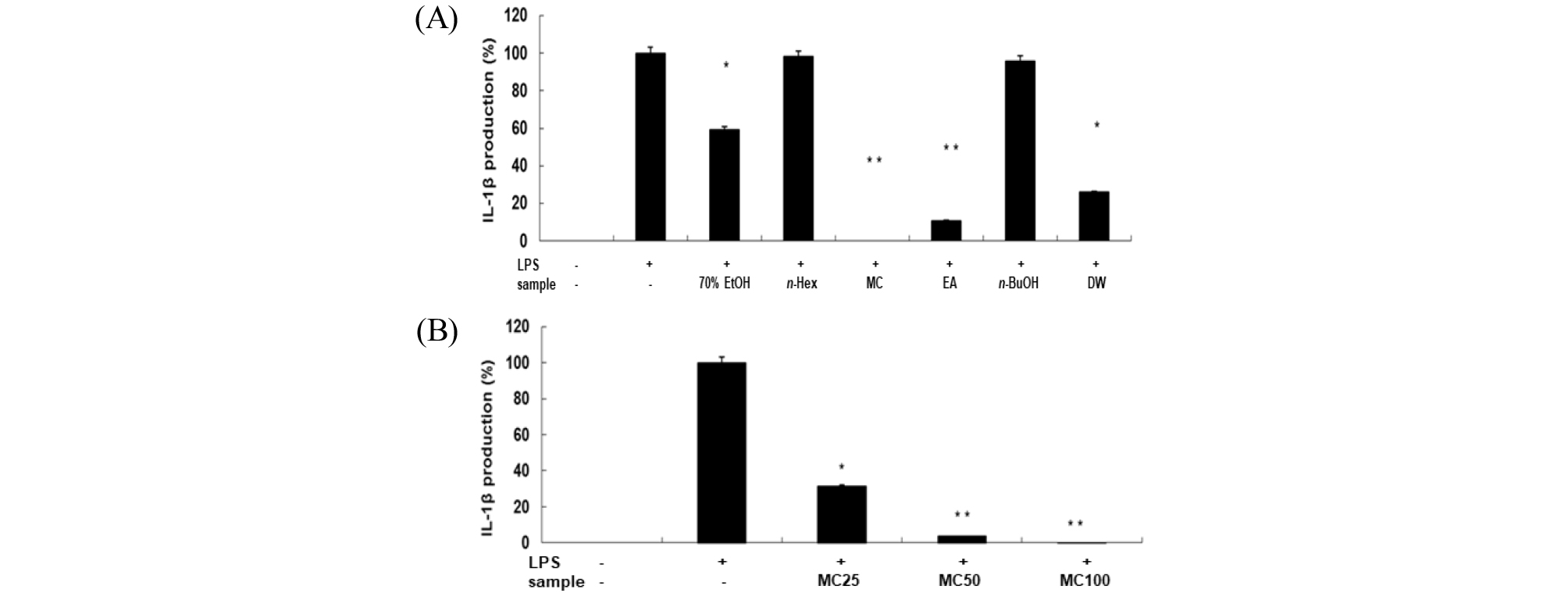
Fig. 6.
Inhibitory effects of crude extract and solvent fractions of P. tenax leaf on IL-1β production in LPS-stimulated RAW 264.7 cells. (A): Cells(1.8 × 105 cells/mL) were treated by LPS(1 ㎍/mL) for 24 h in the presence of crude extract and solvent fractions(100 ㎍/mL) of P. tenax leaf. Supernatants were collected, and the IL-1β concentration in the supernatants was determined by ELISA. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01 (B): Cells were treated by LPS(1 ㎍/mL) for 24 h in the presence of CH2Cl2 (MC) fractions(25, 50 and 100 ㎍/mL).
Chemokine 생성 변화에 미치는 영향
각질형성세포(keratinocyte cell)에서 생성되는 chemokine인 MDC와 TARC는 아토피 피부염의 증상과 밀접한 연관성을 갖고 있다고 알려져 있다(Soumelis et al, 2002). 따라서 TARC와 MDC의 발현을 유도한 HaCaT 세포에서 신서란 잎 조추출물 및 분획물에 의한 TARC와 MDC의 감소량을 측정하여 아토피 억제 효과를 확인할 수 있다(Kwon et al., 2012). 아토피 활성 억제를 확인하기 위하여 human keratinocyte인 HaCaT 세포에 hINF-γ로 자극하여 chemokine 생성을 유도한 후 신서란 추출물과 용매 분획물을 처리하여 그 효과를 조사하였다. hINF-γ무 처리군의 chemokine 생성을 0%, hINF-γ 처리군의 chemokine 생성을 100%로 하여 활성을 조사한 결과는 Fig. 7, 8과 같다. Fig. 6에서 보는바와 같이 methylene chloride와 ethyl acetate 분획물에서 각각 65%, 93%의 MDC 생성 억제효과가 나타났다. 또한, 이들 분획물의 MDC 생성 억제효과는 농도 의존적이었다(Fig. 7B). TARC 생성 억제효과도 methylene chloride와 ethyl acetate 분획물에서는 뚜렷하게 나타났다(Fig. 8).
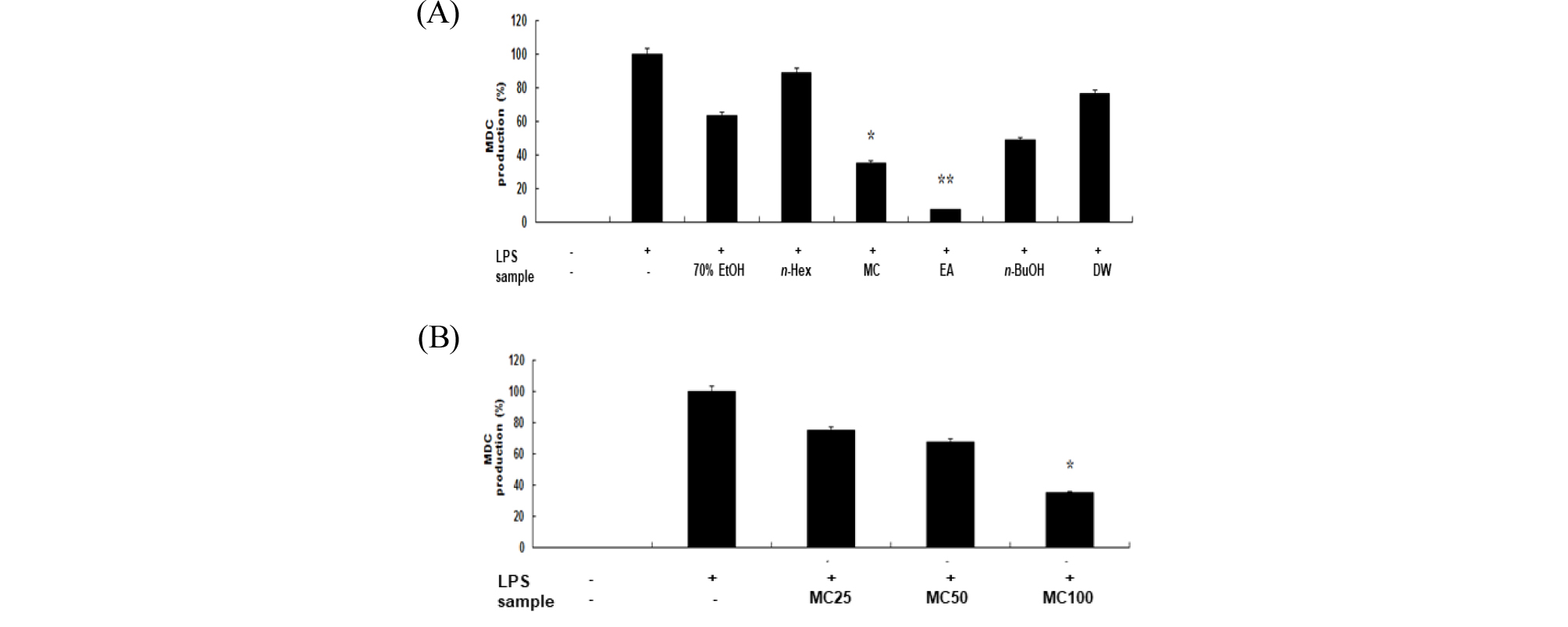
Fig. 7.
Inhibitory effects of crude extract and solvent fractions of P. tenax leaf on MDC production in hIFN-γ-stimulated HaCaT cells. (A): MDC concentration in culture medium was measured with ELISA method in HaCaT cells (3.0 × 105 cells/mL) treated with hIFN-γ (10 ng/mL) in the presence of crude extract and solvent fractions(100 ㎍/mL) of P. tenax leaf. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01. (B) MDC concentration in HaCaT cells (3.0 × 105 cells/mL) treated with hIFN-γ (10 ng/mL) in the presence of CH2Cl2 (MC) fractions (25, 50 and 100 ㎍/mL).
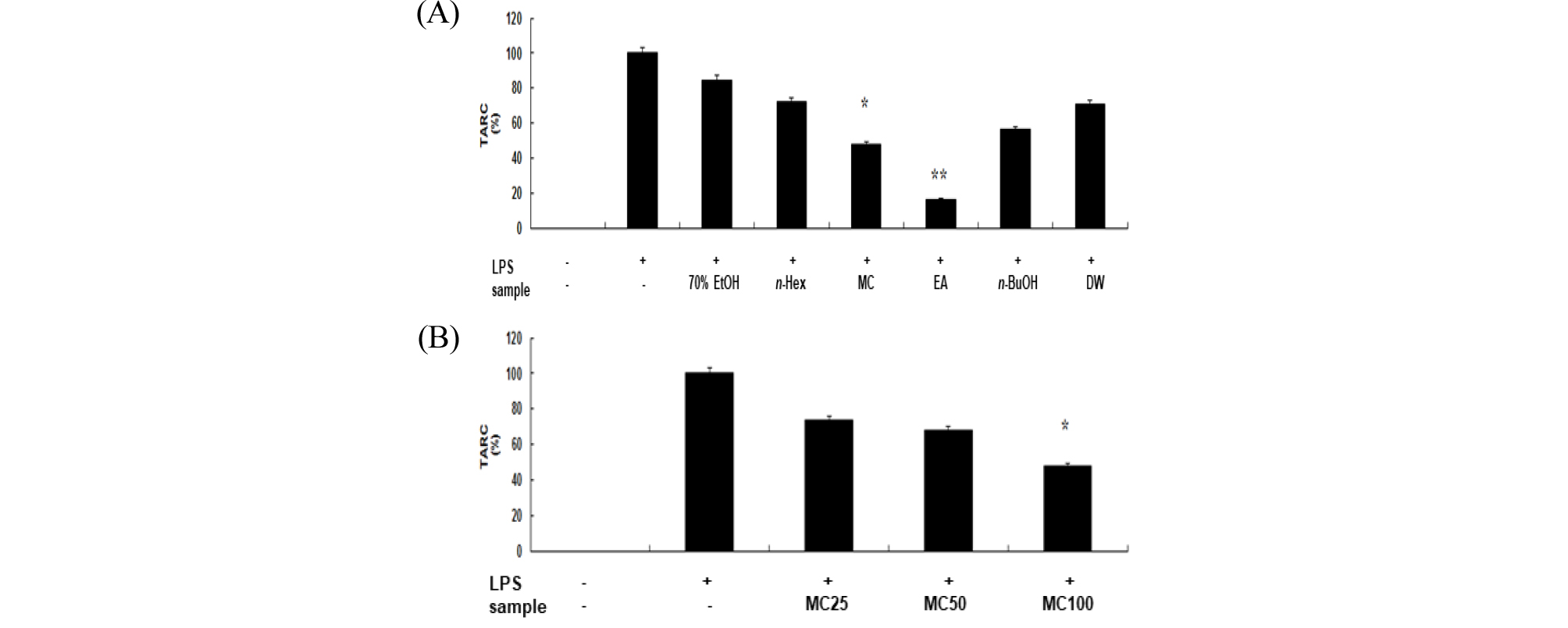
Fig. 8.
Inhibitory effects of crude extract and solvent fractions of P. tenax leaf on TARC production in hIFN-γ-stimulated HaCaT cells. (A): TARC concentration in culture medium was measured with ELISA method in HaCaT cells (3.0 × 105 cells/mL) treated with hIFN-γ (10 ng/mL) in the presence of crude extract and solvent fractions(100 ㎍/mL) of P. tenax leaf. Values are the mean ± SEM of triplicate experiments. *, P<0.05; **, P<0.01. (B) TARC concentration in HaCaT cells (3.0 × 105 cells/mL) treated with hIFN-γ (10 ng/mL) in the presence of CH2Cl2 (MC) fractions (25, 50 and 100 ㎍/mL).
이상의 결과를 종합하여 볼 때 신서란 에탄올 추출물과 용매 분획물은 LPS에 의한 대식세포 염증반응에서 NOS 및 COX-2 발현 억제를 통해 NO, PGE2 생성 억제뿐만 아니라 전염증성 물질인 TNF-α, IL-6, IL-1β의 생성을 억제시켜 항염증 효능이 있음을 제시하고 있다. 또한 아토피 유발 인자인 MDC 및 TARC 생성을 억제시킴으로서 항아토피 효과를 갖는 유효성분이 있음을 보이고 있다. 즉, 신서란 추출물 중 methylene chloride와 ethyl acetate 분획물에 함유된 성분들이 독성이 없고 항염증 및 항아토피 활성을 갖는 주요 분획물임을 제시하는 것이며, 앞으로 이 분획물의 유효성분 물질을 구조적으로 밝히고 기작연구가 진행된다면 항염증 및 항아토피 개선 기능을 가진 기능성 제품의 원료로 사용될 수 있을 것으로 여긴다.
적 요
본 연구는 하라케케(Harakeke)로 불리는 신서란(Phormium tenax)를 화장품 및 의약품산업의 기능성 소재로서의 이용 가능성을 확인하기 위하여 신서란 잎을 대상으로 70% 에탄올 추출물과 용매 분획물을 제조하여, 이것들의 항염증 및 항아토피의 효과를 조사하였다. LPS로 유도된 RAW 264.7 세포에서 신서란 에탄올 추출물과 용매 분획물의 항염증 효과를 조사한 결과, methylene chloride와 ethyl acetate 분획물에서 NO와 PGE2 생성 억제 활성이 가장 높게 나타났으며, 농도 의존적으로 NO와 PGE2 생성 억제 활성을 보였다. 또한, 이들 분획물에서는 iNOS 및 COX-2 발현 억제 활성을 보였다. 신서란 잎 조추출물과 용매 분획물에 의한 NO, PGE2 생성 억제 활성이 NOS 및 COX-2 발현 억제에 의한 것임을 제시한다. 더불어, hIFN-γ로 자극된 HaCaT 세포에 용매 분획물을 처리하여 MDC 및 TRAC 생성억제 효과를 조사한 바, methylene chloride 분획물은 MDC 및 TATC의 생성을 각각 65%, 52% 생성억제 시켰으며, ethyl acetate 분획물은 MDC 및 TATC의 생성을 각각 93%, 84% 억제 효과를 보였다. 이상의 결과는 신서란 잎 조추출물과 용매 분획물을 이용한 항염증 및 항아토피 효능을 갖는 유효성분 분리 및 활용화 연구에 중요한 기초자료가 될 것이며, 기능성 화장품, 의약외품 및 의약품 소재 개발에 적용 가능성이 높다고 사료된다.
