

서 언
국립공원공단에서 수행한 「2019년 지리산국립공원 공원자원조사」에 따르면, 지리산국립공원 내 탐방로는 총 55개 구간으로 총 연장 241.55 ㎞에 달한다. 지난 5년간 지구단위의 탐방객 추이를 보면 2015년부터 꾸준히 증가하고 있으며, 2018년 3,309천 명의 가장 많은 탐방객 수를 기록하였다. 지리산국립공원은 탐방객 이용밀도가 높은 공원 중 하나이며, 탐방편의 관련 요구도 역시 높을 것으로 예상할 수 있다. 반면 이로 인한 자연환경의 훼손 및 교란 위험성과 자연환경에 대한 보전 요구도 역시 높아 질 것으로 판단된다.
산림식생은 산림생태계의 1차 생산자로서 그 지역 토양과 기후특성을 반영하고 있으며, 야생동・식물 서식지로서 핵심 생태축을 담당하고 있다(Byeon and Yun, 2017; Jang et al., 2008; Shin and Yun, 2014). 식생에 의한 산림분류 및 특성파악은 산림의 보전 및 관리를 위한 객관적・생태적 관리단위를 제공하기 때문에(Bae et al., 2003; Byeon and Yun, 2016; 2017; Kim and Yun, 2009; Lee and Yun, 2002; Yun et al., 2007), 산악형 국립공원의 보전・관리를 위해 식생 조사 및 특성파악은 반드시 필요한 부분이라 할 수 있다.
지리산국립공원은 우리나라 남부지방의 내륙 한가운데 위치하고 해발고도 110 ∼ 1,915 m의 표고차를 보이는 등 환경조건이 다양하여 식물상 및 분포에 대한 조사가 일찍부터 이루어졌다(Gwon et al., 2013). 1900년대 초 일본학자(Hatusima, 1934)에 의해 식물상에 대한 초기연구가 진행되었으며, 이후 주요 지역별 군락구조 및 천이과정 등 식생학적 연구(Jung et al., 1996; Kang, 1984; Kim, 1975; Kim, 1988; Lee and Kim, 1999; Park et al., 1991; Park et al., 2003; Yim and Kim, 1992)로 발전되었다. 이밖에 지리산국립공원 훼손지 내 식생회복 및 복원에 관한 연구(Jung et al., 2001; Kwon et al., 1991; Oh et al., 1997; Oh, 2017; Park, 2009; Shin et al., 2013)도 수행된 바 있다. 이처럼 지리산국립공원은 수많은 연구자들이 다양한 주제로 관심을 갖고 연구하고 있는 대표적인 동・식물 서식지 중 하나이다.
자연공원법 제17조의3에 따라 국립공원을 포함한 자연공원은 매 10년마다 보전・관리계획을 수립하여야 하며, 동법 시행령 제13조의2에 따라 자연생태 등 해당 공원의 특성을 최대한 반영하도록 명시하고 있다. 지리산국립공원은 지난 2012년 제1차 보전・관리계획(2013 ∼ 2022년)이 수립된 바 있으며, 2022년까지는 제2차 공원별 보전・관리계획(2023 ∼ 2032년)의 수립이 필요한 상황이다. 이를 위한 기초자료로서 지리산국립공원 전반에 대한 산림식생의 최신 정보 및 자료 축적이 필요한 실정이다.
또한 제4기 국립공원 공원자원조사는 기존과 다르게 동일한 정점(각 공원별 위치와 개수가 정해짐)에서 다양한 동‧식물 분류군 조사가 동시에 이뤄졌다. 즉, 모든 분류군(식생, 식물상, 포유류, 조류, 파충류, 곤충류)조사가 동일 한 공간범위 내에서 수행되며, 이를 바탕으로 각 정점별 서식지 평가가 이뤄질 수 있도록 하였다. 그러나 식생을 제외하면 대부분 시기별 출현 종 조사에 국한되어 있어 서식지 평가를 위한 입지‧환경 정보가 부족한 실정이다.
이에 본 연구는 지리산국립공원 전체를 대상으로 표본지 매목조사를 통한 식물군집구조를 파악하고자 하였으며, 제2차 지리산국립공원 보전・관리 계획 및 제4기 지리산국립공원 공원자원조사 서식지 종합평가를 위한 기초자료 수집 및 제공을 목적으로 수행되었다.
재료 및 방법
조사대상지 개황
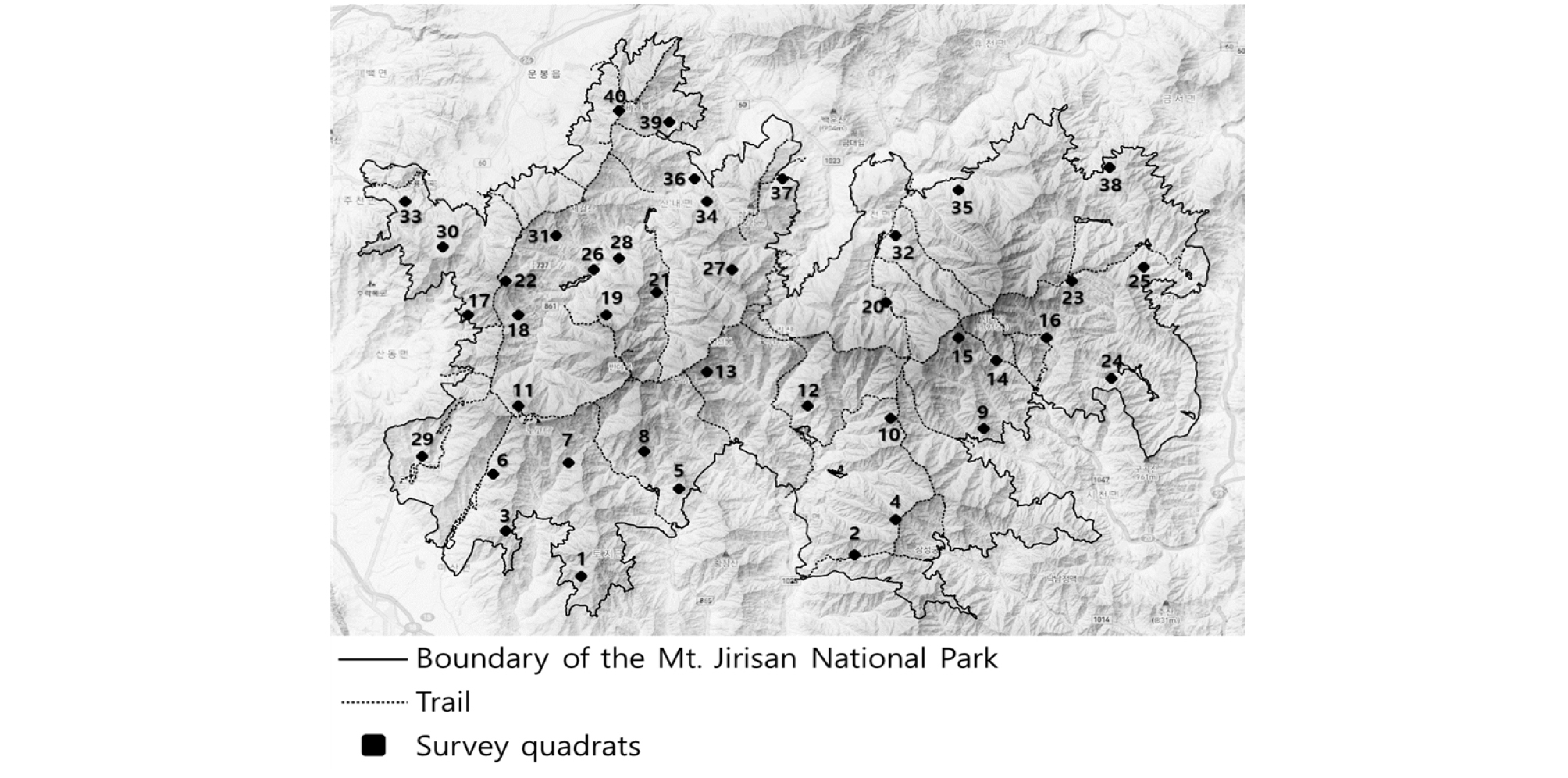
지리산국립공원은 북위 35°12′45.23″∼ 35°26′52.31″, 동경 127°27′11.44″∼ 127°49′32.01″사이에 위치(WGS 84 기준)해 있다. 1967년 12월 29일 지정된 우리나라 최초의 국립공원으로, 총 면적은 483.022 ㎢이다(Fig. 1). 전국 22개 국립공원 중 육상면적이 가장 넓은 산악형 국립공원으로서, 경상남도 산청군・하동군・함양군, 전라남도 구례군, 전라북도 남원시 등 3개의 도, 1개의 시, 4개의 군, 15개의 읍・면에 걸쳐있다. 천왕봉(1,915 m), 반야봉(1,732 m), 노고단(1,507 m)을 중심으로 다수의 봉우리가 분포하고 있다.
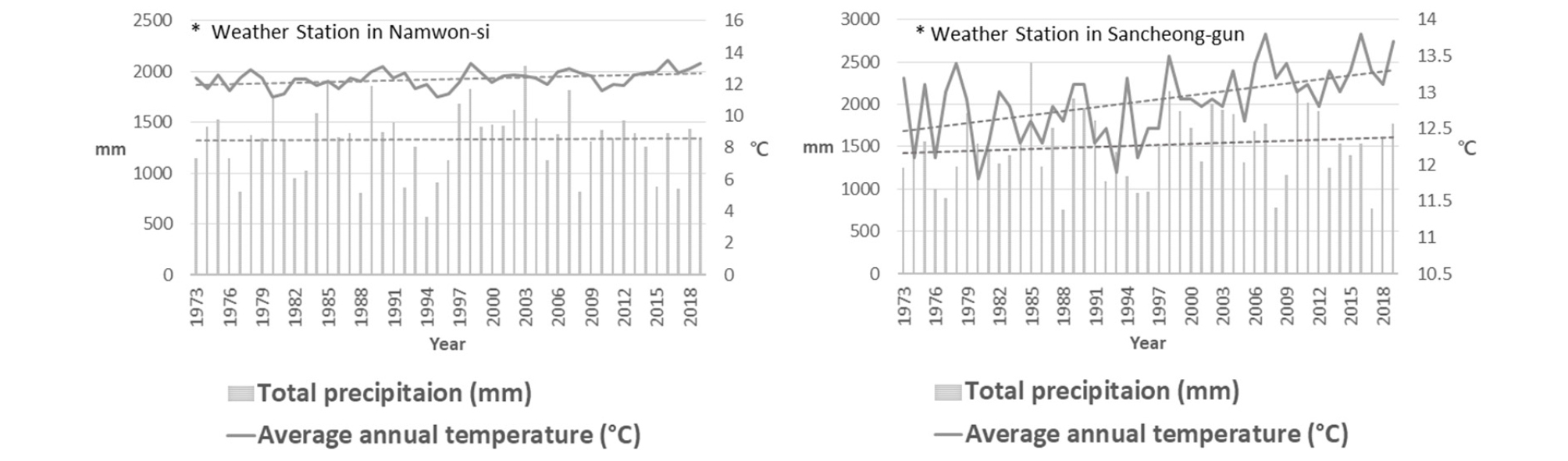
기상청 기상자료개방포털(https://data.kma.go.kr/)에 따른 지리산국립공원 일원 남원과 산청의 지난 47년 간 연 평균 기온은 각각 12.3℃, 12.9℃로 나타났으며, 연 평균 총 강수량은 각각 1,334.7 ㎜, 1,516.4 ㎜로 확인되었다. 두 지역의 연 평균 기온 및 연간 총 강수량 모두 증가 경향을 보이고 있으며, 산청 지역이 남원 지역에 비해 증가폭이 높은 것으로 확인되었다(Fig. 2).
조사방법
표본지 조사
식물군락의 구조를 조사하기 위한 표본조사법으로 방형구법이 일반적으로 사용되며(Brower and Zar, 1977), 방형구법에 있어서 표본추출 방법은 최소면적의 단일 조사구를 설치하거나 몇 개의 소조사구를 설치하는 것이 일반적인 방법이다(Park and Seo, 2002). 본 조사는 국립공원공단에서 수행한 제4기 지리산국립공원 공원자원조사에서 정해진 40개의 정점(모든 동‧식물 분류군이 동일한 정점을 기준으로 조사 수행)을 기준으로 수행되었으며, 당해 정점을 기준으로 주변에 임분이 균질하고 인위적인 교란이 없는 식생을 대상으로 방형구법에 의한 표본조사를 수행하였다. 식물군락의 구분은 상관식생을 기준하였으며, 조사기간은 2019년 6월부터 11월까지이다.
기본적인 조사방법은 국립공원공단에서 정한 제4기 공원자원조사 식생 분야 매뉴얼을 따랐다. 표본지 내 20 m × 20 m 크기의 정방형구 형태의 조사구 40개(16,000 ㎡)를 구축하였으며, 층위별 중첩방형구법에 따라 조사하였다. 교목층은 20 m × 20 m (전체 면적), 아교목층은 10m × 10 m(교목층 면적의 1/4), 관목층은 5 m × 5 m(아교목층의 1/4) 면적범위 내에서 출현하는 모든 개체를 조사하였다. 단, 아교목층과 관목층은 대표성을 나타내는 사분면을 선택하여 조사하였다. 층위구분을 위한 수고기준은 교목층 8 m 이상, 아교목층 2 ∼ 8 m, 관목층 2 m 이하로 하되, 해당 조사구의 고유한 식생구조와 종에 따라 조사자가 판단하여 가감하여 정하였다. 초본층은 교목층과 동일한 면적(20 m × 20 m) 범위에서 조사를 수행하였으며, 종별 평균 초장과 피도를 조사하였다. 목본의 유묘 혹은 치수의 경우, 수고에 관계없이 관목층에 기입하였으며, 초본 역시 초장에 관계없이 초본층에 기입하였다. 조릿대 등의 대나무류는 높이에 관계없이 초본층에 기입하였다.
분류군의 확인 및 동정은 Kim and Kim (2001), Lee (1996), Lee (2003)의 도감을 이용하였고, 양치식물은 Korean Fern Society (2005)의 문헌을 참고하였다. 학명과 국명은 국립수목원과 한국식물분류학회가 제시한 국가표준식물목록(Korea National Arboretum and The Korean Society of Plant Taxonomists, 2007)에 준하였다.
자료분석
상대우점치 및 종다양성지수 분석
층위별 각 수종의 상대적 우세도를 비교하기 위해 Curtis and McIntosh (1951)의 중요도를 백분율로 표현한 상대우점치(IP, Importance Percentage; Brower and Zar, 1977)를 분석하였으며, 층위별 가중치를 부여하고 군락 내 모든 종의 우세도를 비교하기 위해 평균상대우점치(MIP, Mean Importance Percentage; Yim and Cheon, 1980)를 분석하였다. 우점치 분석은 산림군집 내 각 식물종의 영향력과 우세력을 평가하기 위한 방법으로 산림의 종구성을 객관적으로 파악하기에 용이하다(Brower and Zar, 1977; Byeon and Yun, 2017).
또한, 종 구성 상태의 다양한 정도를 확인하기 위해 Shannon의 수식(Pielou, 1975; Shannon and Weaver, 1963)을 적용하여, 종 다양도, 최대 종 다양도, 균재도, 우점도를 분석하였다. 종다양성지수는 종수와 개체수라는 제한된 변수만으로 산출된다는 한계가 있지만(Byeon and Yun, 2017; Krebs, 1985), 산림식생의 안전성을 유추할 수 있다(Byeon and Yun, 2017). 또한 서로 다른 지역의 종 다양성을 비교할 수 있다는 장점을 가지고 있다(Byeon and Yun, 2017; Park et al., 2001). 본 조사에서는 초본층을 제외한 목본식물을 대상으로 하였다.
Classification 및 Ordination 분석
지점별 식생조사 자료를 바탕으로 군락별 군락구조 특성을 파악하기 위해 TWINSPAN (Hill, 1979a)에 의한 군락분류를 실시하였다. TWINSPAN은 계층적 군화분석 방법으로 일차적으로 출현한 생물 종을 기반으로 표본단위를 먼저 분류하고, 이를 기반으로 다시 출현생물을 분류함에 따라 다른 분류방법에 비해 환경요인의 선호에 따른 생물 특징을 구분하기 용이한 방법이다(Hill, 1979a; Min et al., 2018). TWINSPAN의 통계학적 신뢰성은 선행연구(Cao et al., 1997; Gauch Jr and Whitaker, 1981)를 통해 검증된 바 있으며, 생태학 분야에서 생물의 군집과 환경요인 관계를 분석하고자 할 때 TWINSPAN이 주요 이용되고 있다(Brown and May, 2000; Kazanci et al., 2008; Lee et al., 2000; Min et al., 2018).
군락분류를 통해 그룹화된 군락의 분포특성을 파악하기 위해 DCA (detrended correspondence analysis) ordination (Hill, 1979b) 분석을 실시하였다. Ordination 기법은 군락구조를 밝히고 군락 내 식생과 입지환경과의 상호작용을 판단하기 위한 방법으로 알려져 있다(Byeon and Yun, 2017, Byeon et al., 2020; Greigh-Smith, 1983; Ter Braak, 1988; Yun and Hong, 2000). 특히 DCA 기법은 초기 ordination 기법이었던 PO (polar ordination)과 이를 개선한 RA (reciprocal averaging) 및 PCA (principal component analysis)의 단점을 보완한 기법으로, 제2축이 종종 제1축에 종속되고(arch effect) 축 양극단 샘플들의 변이가 중앙의 샘플들보다 훨씬 적게 나타나는 왜곡현상(edge effect)을 개선한 기법이다(Hill, 1979b; Ri, 1994). 또한 DCA는 환경요인과의 관계성을 분석할 수 있어, 식생구조의 정량적 분석을 통한 군집생태학 연구에 널리 사용되고 있다(Palmer, 2004).
본 조사에서는 초본층을 제외한 목본식물을 대상으로 각 조사구별 평균상대우점치와 입지환경 자료를 바탕으로 Classification과 Ordination을 실시하였으며, PC-ORD 6 소프트웨어를 사용하였다. 또한 군락분류를 통해 그룹화된 군락 간 종 구성의 유사성을 비교하기 위해 유사도지수(Brower and Zar, 1977)를 추가 분석하였다.
결 과
조사구별 입지환경 및 출현종
조사구별 해발고도는 425 m부터 1,711 m까지 분포하며, 경사도는 10°부터 45°까지 분포하였다. 토양은 모두 갈색삼림토였으며, 사면과 평면 지형에 위치한 식생군락이 주요한 것으로 나타났다. 낙엽부식층은 10 ㎝ 미만으로 나타났으며, 암석노출비율은 대부분 50% 미만으로 확인되었다. 식생보전등급 Ⅱ등급에 해당하는 식물군락이 주요했으며, 대부분 중용에 해당하는 수분조건을 나타냈다. 우점종 영급은 대부분 6영급 이상인 것으로 확인되었다. 각 조사구별 자세한 입지환경 정보는 Appendix 1, 각 조사구별 식생구조 정보는 Appendix 2와 같다.
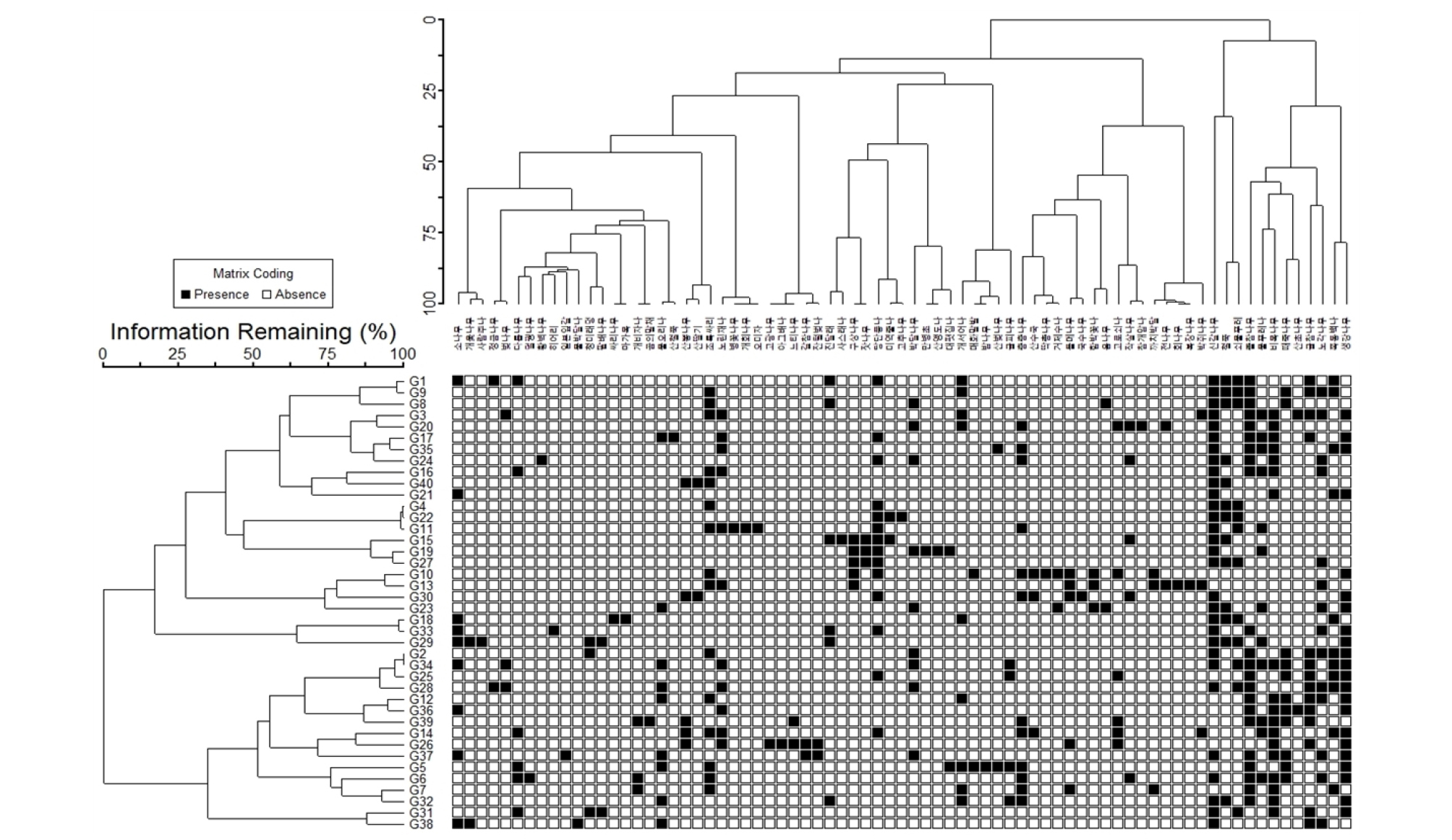
각 조사구와 조사구 내 목본식물의 출현종에 대한 관계성 파악을 위해 Two-way Cluster 분석을 실시하였다. 주로 신갈나무(30개소), 생강나무(22개소), 졸참나무(22개소), 비목나무(18개소), 굴참나무(14개소), 노각나무(14개소), 당단풍나무(14개소), 철쭉(13개소), 물푸레나무(12개소), 쇠물푸레(12개소), 층층나무(12개소), 노린재나무(11개소), 때죽나무(11개소), 쪽동백나무(11개소)가 10개 이상의 조사구에서 다수 출현하였다. 반면, 개회나무, 고광나무, 고추나무, 곰의말채나무, 국수나무, 덜꿩나무, 딱총나무, 마가목, 만병초, 물박달나무, 밤나무, 병꽃나무, 복장나무, 사람주나무, 사스래나무, 산앵도나무, 산철쭉, 싸리, 아그배나무, 오미자, 일본잎갈나무, 참개암나무, 황벽나무, 회나무, 히어리는 단 1개의 조사구에서만 희소하게 출현하는 것으로 확인되었다(Fig. 3).
Classification
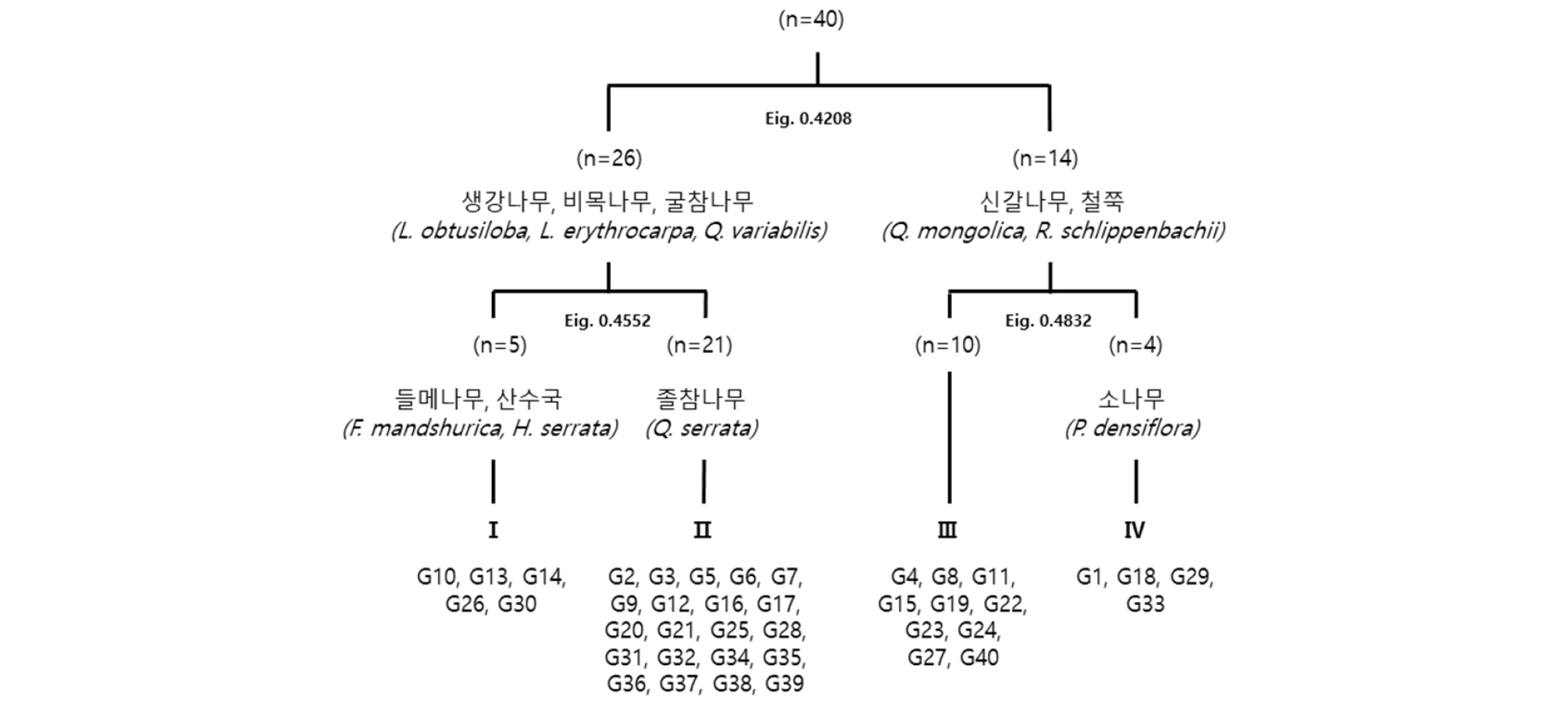
전체 40개 조사구에 대한 유형별 군락분류결과 첫 번째 단계(level 1)에서 생강나무(Lindera obtusiloba Blume)・비목나무(Lindera erythrocarpa Makino)・굴참나무(Quercus variabilis Blume) 그리고 신갈나무(Quercus mongolica Fisch. ex Ledeb.)・철쭉(Rhododendron schlippenbachii Maxim.)에 의해 2개의 그룹으로 유형분류 되었으며, 두 번째 단계(level 2)에서 1그룹은 들메나무(Fraxinus mandshurica Rupr.)・산수국(Hydrangea serrata f. acuminata (Siebold & Zucc.) E.H.Wilson) 그리고 졸참나무(Quercus serrata Thunb. ex Murray)에 의해 2개의 군락, 2그룹은 소나무(Pinus densiflora Siebold & Zucc.)에 의해 2개의 군락으로 유형화 되었다. 총 4개 군락으로 Ⅰ군락은 들메-당단풍나무군락(Fraxinus mandshurica-Acer pseudosieboldianum Community), Ⅱ군락은 졸참-굴참나무군락(Quercus serrata-Quercus variabilis Community), Ⅲ군락은 신갈나무군락(Quercus mongolica Community), Ⅳ군락은 소나무군락(Pinus densiflora Community)으로 분류되었다(Fig. 4).
Ordination 및 군락구조
TWINSPAN 기법으로 유형화된 4개의 군락을 기준으로 그래프를 작성하였으며(Fig. 5), 제1축(0.61955)과 제2축(0.46558)의 eigenvalue가 높게 나타나 x축에 제1축, y축에 제2축을 지정하여 모든 군락을 배치하였다. 4개의 군락 모두 제1축과 제2축을 기준으로 불연속적인 분포를 보이며 각기 구분되는 모습을 볼 수 있다. 다만 Ⅲ군락에 속한 G23번 조사구가 분리되어 나타났으며, Ⅰ군락에 가깝에 분포하는 것을 볼 수 있다. G23번 조사구는 신갈나무 우점군락으로 신갈나무군락으로 유형화된 Ⅲ군락에 포함되었지만, 동반종에 의해 Ⅰ군락과의 유사성이 높은 것으로 판단된다.
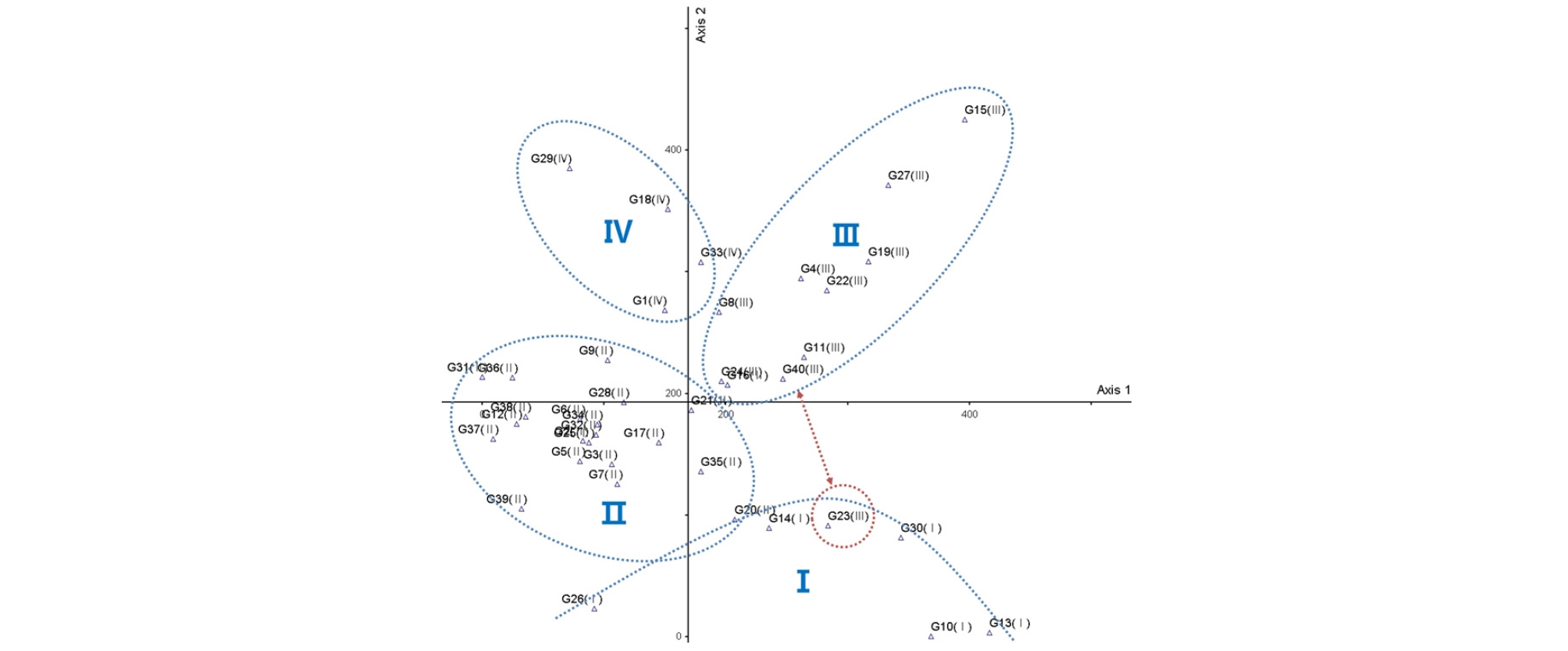
Ⅰ군락은 들메나무군락, Ⅱ군락은 졸참-굴참나무군락, Ⅲ군락은 신갈나무군락, Ⅳ군락은 소나무군락으로 구분되었다. 각 군락별 주요 목본 수종의 층위별 상대우점치와 평균상대우점치는 Table 1과 같으며, 본 조사의 모든 목본 수종에 대한 목록과 이를 통합한 층위별 상대우점치, 평균상대우점치는 Appendix 3과 같다.
Table 1.
Importance Percentage of woody species by stratum in each community
DCA ordination에 따른 주요 수종의 우점도별 조사구 분포 경향성은 다음과 같다(Fig. 6). Ⅰ군락 우점종인 들메나무는 제1축에 의해 동일 군락 내 다른 조사구와 구분(r=0.485 tau=0.229)되었다. Ⅱ군락 우점종인 졸참나무는 기존 Ⅱ군락의 분포범위와 유사하게 나타났으며, 제1축에 의한 설명력(r=-0.472 tau=-0.396)이 상대적으로 높게 나타났다. 또한 Ⅱ군락에 인접하고 타 군락에 속한 일부 조사구에서도 높은 우점도를 나타냈다. Ⅲ군락 우점종인 신갈나무는 제1축에 의해 Ⅲ군락에 속한 다른 조사구와 구분(r=0.443 tau=0.374)되었으며, Ⅲ군락에 인접하고 타 군락에 속한 일부 조사구에서 높은 우점도를 나타냈다. Ⅳ군락 우점종인 소나무는 기존 Ⅳ군락에 속한 조사구의 분포와 일치하였으며, 제2축에 의한 설명력(r=0.469 tau=0.305)이 상대적으로 높게 나타났다.
TWINSPAN 군락분류 결과에 따른 군락별 종다양성지수 분석결과는 다음과 같다(Table 2). Ⅰ군락과 Ⅱ군락, Ⅱ군락과 Ⅳ군락이 50% 이상의 유사도지수 값을 나타내, 각 군락의 출현종수 대비 공통으로 출현한 종이 상대적으로 많은 것으로 나타났다. 반면 Ⅰ군락과 Ⅳ군락이 가장 낮은 유사도를 나타내, 각 군락의 출현종수 대비 공통으로 출현한 종이 상대적으로 적은 것으로 나타났다(Table 3).
Table 2.
Species diversity index by community
Table 3.
Similarity index by community
| Community | Ⅰ | Ⅱ | Ⅲ |
| Ⅱ | 53.66% | ||
| Ⅲ | 41.79% | 45.78% | |
| Ⅳ | 29.09% | 50.70% | 32.14% |
TWINSPAN 군락분류 결과에 따른 군락별 입지환경의 산술적 차이는 다음과 같다(Table 4). Ⅲ군락의 해발고도 분포가 가장 높은 것으로 나타났으며, Ⅲ과 Ⅳ군락의 경사도가 Ⅰ과 Ⅱ군락에 비해 높은 것으로 확인되었다. 낙엽부식층의 차이는 미미했으며, Ⅰ군락의 암석노출비율이 평균 51% 수준으로 다른 군락에 비해 높게 나타난 반면, Ⅳ군락은 평균 6% 수준으로 암석노출이 거의 없는 것으로 확인되었다.
Table 4.
Environmental factor by community
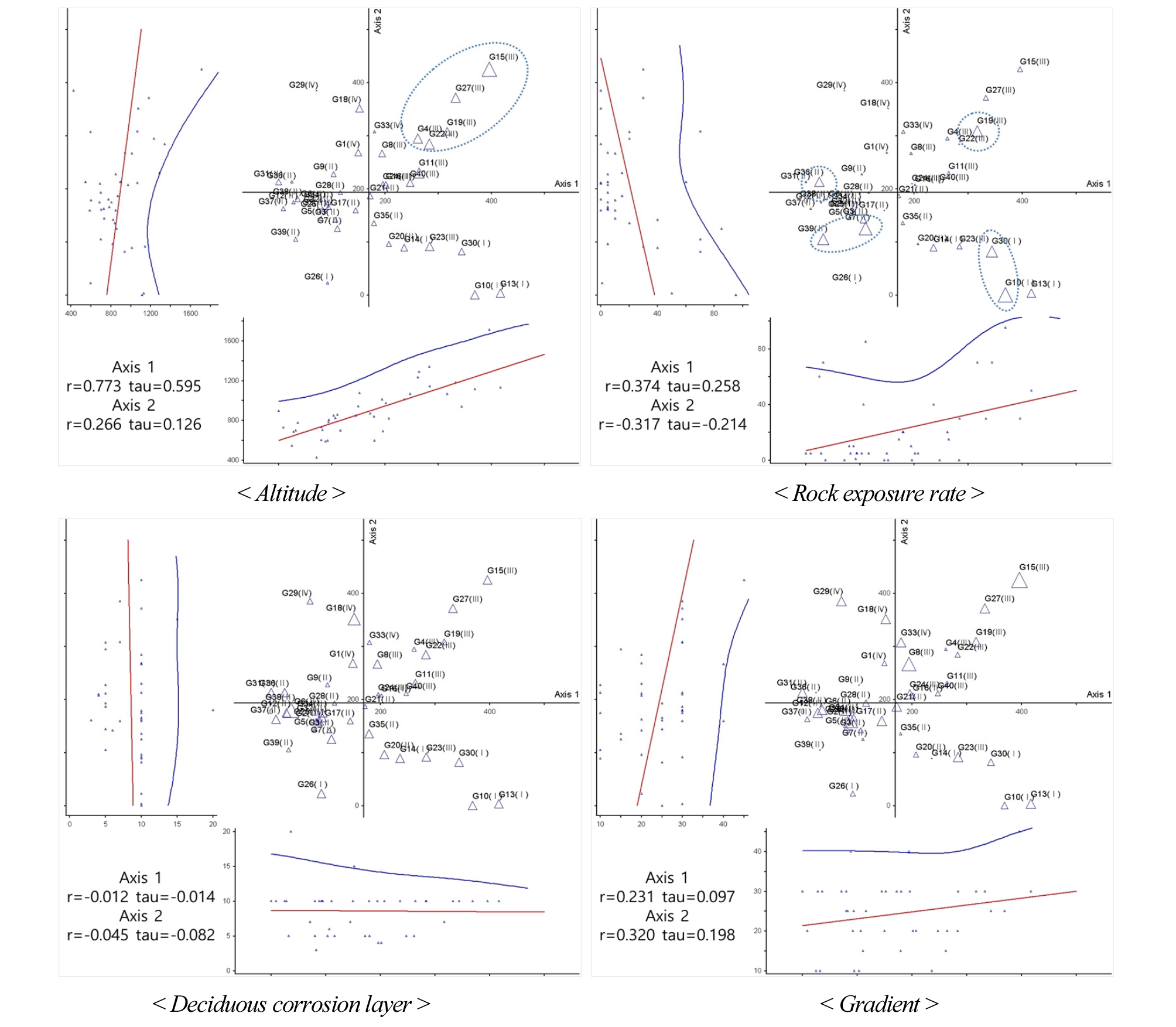
DCA ordination에 따른 입지환경별 조사구 분포 경향성은 다음과 같다(Fig. 7). 해발고도의 경우 Ⅲ군락이 높은 것으로 나타났으며, 제1축에 의해 Ⅲ군락 내 몇몇 조사구가 상대적으로 높은 설명력(r=0.773, tau=0.595)으로 구분되었다. 암석노출비율은 Ⅳ군락을 제외한 모든 군락 내 몇몇 조사구에서 높게 나타났으며, 제1축과 제2축 모두 경향성 없이 산재하여 분포하는 것으로 나타났다. 낙엽부식층과 경사도는 두 축에서 모두 경향성이 없는 것으로 나타나 군락별 큰 차이가 없는 것으로 나타났다.
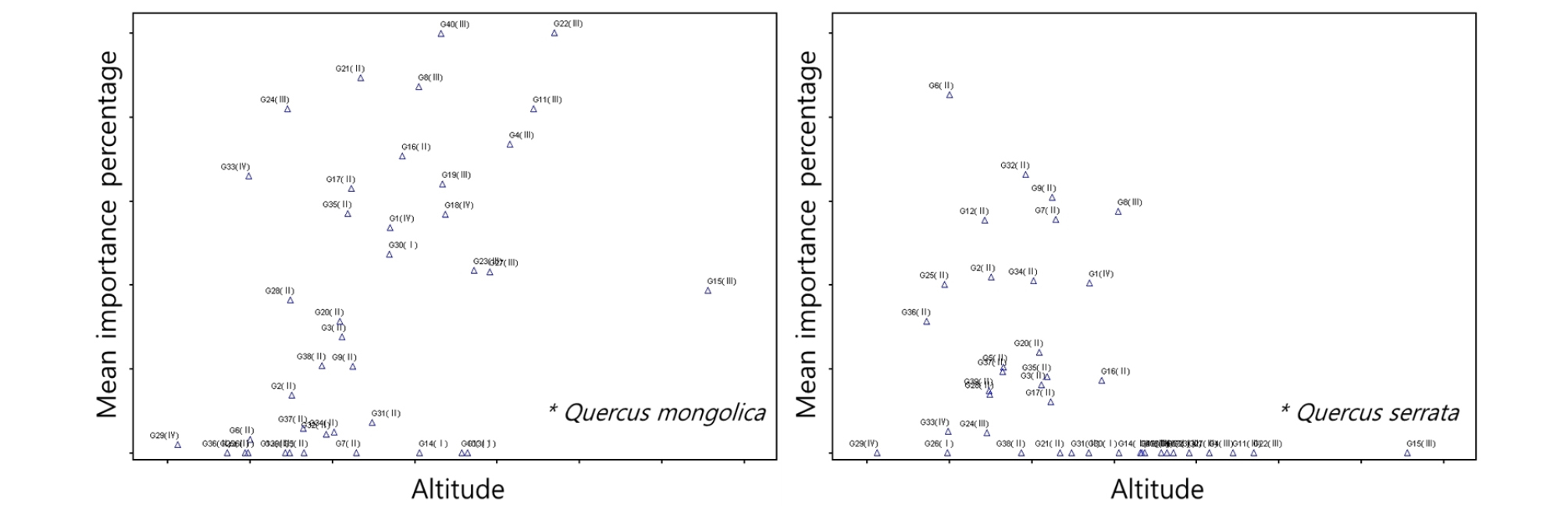
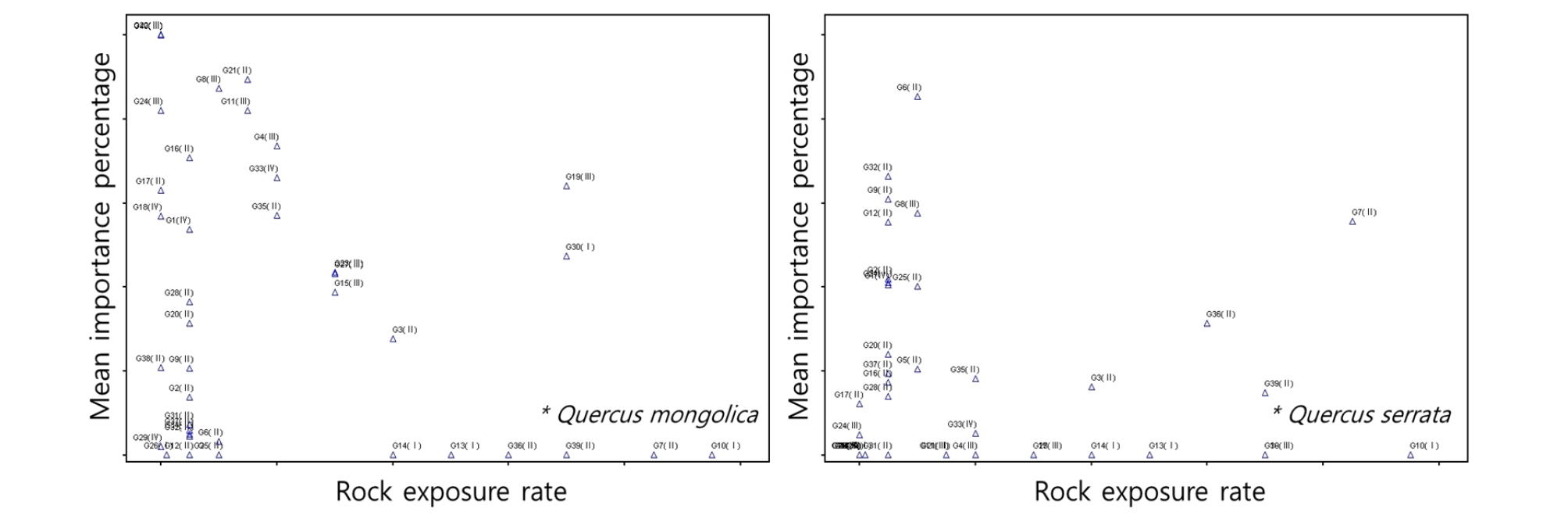
해발고도에 따른 신갈나무 우점도는 500 ∼ 1,400 m 사이에서 상대적으로 높은 우점도를 나타냈으며, 졸참나무와 굴참나무는 전반적으로 모든 해발고도에서 낮은 수준의 우점도를 보였지만, 1,000 m 미만에서는 상대적으로 높은 우점도를 보이는 조사구가 분포하였다(Fig. 8). 암석노출에 따른 신갈나무, 굴참나무, 졸참나무의 우점도는 0 ∼ 20% 구간에서 상대적으로 높은 우점도를 보이는 조사구가 다수 분포하였다(Fig. 9). 소나무는 전반적으로 암석노출비율에 관계없이 낮은 우점도를 보였으며, 일부 암석노출비율이 낮은 구간에서 높은 우점도를 보인 조사구가 확인되었다. 기타 경사도, 낙엽부식층, 보전등급에 따른 주요종의 분포특성은 나타나지 않았다. 이들 항목은 데이터 범위가 상대적으로 좁고, 세분화되어 있지 않기 때문으로 판단된다. 또한 Ⅰ군락에서 최우점하고 있는 들메나무의 경우 모든 입지환경 측정항목에서 특이성이 없었으며, 대부분 낮은 수준의 우점도를 보이는 조사구가 다수 분포하고 있다.
고 찰
40개 표본지 내 주요 우점종은 신갈나무, 졸참나무, 굴참나무, 박달나무, 소나무, 들메나무, 느티나무, 층층나무, 개서어나무, 구상나무로 파악되며, 앞서 언급한 선행연구(Gwon et al., 2013; Park et al., 1991; Park et al., 2003; Yim and Kim, 1992)에서 확인된 군락의 범위를 벗어나지 않는 수준으로 판단된다. 특히, 굴참나무, 졸참나무, 신갈나무, 소나무, 들메나무가 주요하게 출현하였으며, 타 수종들과 상대적으로 높은 빈도로 결합하고 있는 것으로 판단된다. 조사구별 목본식물의 출현종 수는 4종부터 15종까지 조사되었으며, 출현종 수가 낮을수록 종다양도 역시 낮은 값을 나타내고 있었다. 이는 최대종다양도가 종수에 따라 결정되기 때문으로 판단되지만, 단순히 종다양도의 수치적 높고 낮음을 비교하기보다 최대종다양도 대비 종다양도의 비율을 따져보는 것이 합리적일 것이다.
군락분류결과에 따른 군락별 종다양성지수 분석결과, 전반적으로 모든 군락(Ⅰ ∼ Ⅳ)이 최대종다양도 대비 높은 종다양도와 균재도 값을 나타내고 있어, 양호한 종다양성과 출현종별 고른 개체수 분포를 나타내고 있는 것으로 판단된다. 특히 Ⅲ군락(신갈나무군락)과 Ⅳ군락(소나무군락)의 경우, 서어나무 혹은 졸참나무로 천이하는 과정에서 종다양성이 증가한다는 Choi (1983)과 Park et al. (1991)의 선행연구결과가 있다. 그러나 각 군락별 상대우점치 분석결과에 따른 수종 간 경쟁관계를 살펴보면, 아직 서어나무나 졸참나무로의 천이를 판단하기 어려운 것으로 판단된다. 다만, Ⅳ군락(소나무군락) 내 신갈나무의 상층과 중층의 상대우점치가 타 수종에 비해 높게 나타나고 있어, 소나무의 쇠퇴 이후 신갈나무로의 천이가 예상된다. 기타 특이사항으로 Ⅳ군락(소나무군락)이 22종으로 타 군락에 비해 가장 적은 종수를 나타냈으며 균재도 역시 상대적으로 낮게 나타나, 다른 군락에 비해 특정 출현종의 개체수가 편중되어 있고, Ⅰ군락(들메나무군락)은 종수에 비해 개체수가 적은 편이었지만, 균재도가 가장 높아 상대적으로 다른 군락에 비해 출현종별 개체수 분포가 상대적으로 균등한 것으로 판단된다.
Ⅰ군락은 들메나무군락으로 교목층에 들메나무가 우점하고 있으며, 층층나무와 신갈나무가 경쟁하며 세력을 유지하고 있다. 지리산 내 들메나무군락의 존재는 An et al. (2010), Choo et al. (2009), Kim et al. (1991) 등 다수의 선행연구에서 확인되고 있으며, 들메나무와 층층나무가 경쟁 관계에 있다는 정보는 Gwon et al. (2013)에서 확인할 수 있다. 다만, 신갈나무를 극상수종으로 정리한 Yim and Kim (1992)의 연구결과를 비춰보면, 지리산에서는 신갈나무의 세력권 확장이 층층나무에 비해 우세할 것으로 판단되므로 향후 잠재자연식생은 신갈나무군락이 될 것으로 예상된다.
Ⅱ군락은 졸참-굴참나무군락으로 교목층에 굴참나무와 졸참나무가 경쟁하며 우점하고 있으며, 아교목층에서 쪽동백나무와 때죽나무가 경쟁하며 우점하고 있다. 관목층에서는 생강나무가 우점하고 있으며, 아교목성 수종인 비목나무와 관목성 수종인 개비자나무가 경쟁하며 세력을 확장하고 있다. 신갈나무와 함께 극상수종으로 인식되고 있는 졸참나무(Song, 2008)의 세력확장을 확인할 수 있는 사례로 판단된다.
Ⅲ군락은 신갈나무군락으로 신갈나무가 교목층에서 67%, 아교목층에서 27%로 높은 우점도를 보이며, 당단풍나무가 아교목층에서 신갈나무와 경쟁하고 있다. 관목층에서 철쭉, 진달래, 노린재나무가 경쟁하며 하층을 점유하고 있다. 신갈나무군락의 지리산을 대표하는 군락 중 하나로 거의 모든 선행연구에서 다루고 있으며, 높은 우점치와 당단풍나무와 결합하여 군락을 이루고 있다는 점에서 Yim and Kim (1992)의 연구와 맥을 같이 하고 있다.
Ⅳ군락은 소나무군락으로 교목층에서 소나무가 74%의 높은 우점도를 보였다. 아교목층은 신갈나무가 우점하고 있으며, 개서어나무와 쇠물푸레가 경쟁하며 동반발달하고 있다. 관목층에서는 철쭉이 43%로 높은 우점도를 보이고 있으며, 교목성 수종인 물푸레나무와 쇠물푸레의 세력이 확인되었다. 철쭉, 진달래 등과 결합하여 군락을 이룬다는 과거 조사결과Yim and Kim (1992)와는 달리 신갈나무의 세력확장을 확인할 수 있었으며, 향후 소나무의 쇠퇴 이후 신갈나무로의 천이가 예상된다. 지리산의 신갈나무를 기후적 극상림으로 해석하고 있는 Yim and Kim (1992)의 연구결과와 맥을 같이 한다.
Ordination 기법에 의한 분석에서 구분된 각각의 조사구들은 점진적이고 불규칙한 배열에도 불구하고 군락분류에 따른 4가지 군락으로 구별되어 묶여질 수 있으며, 종 조성의 점진적인 차이에 기초한 군락의 구배는 환경요인의 구배와 결합되어 분리되는 것으로 보아, 분리된 각 군락들은 서로 특징적인 환경구배에서 분포하고 있다고 판단할 수 있다.
지리산국립공원 자연자원의 보전 및 관리를 위해 조릿대의 생육 및 번식 관련 모니터링 및 연구가 필요할 것으로 판단된다. 본 조사에서 총 40개 조사구 중 24개 조사구에서 조릿대가 밀생(식피율 50% 이상)하고 있는 것으로 조사되었기 때문이다. 조릿대의 밀생은 식생의 기반이 되는 토양특성을 일률적으로 변화시킬 가능성이 높으며, 갱신치수를 현저하게 피압하여 고사시키고, 타감작용으로 다른 종자의 발아를 억제(Li et al., 1992; Nakashizuka, 1984; Peter et al., 1992; Park et al., 2012)하는 등 식물의 종다양성을 저하시키는 동시에 천연갱신에 큰 부담을 주는 것으로(Iwamoto and Sano, 1998; Lei and Koike, 1998; Nakashizuka, 1984; Park et al., 2012; Yuruki et al., 1977) 알려져 있다. 이에 따라 조릿대가 밀생하고 있는 산림지역에 대한 모니터링을 통해 조릿대가 주변 식생천이에 미치는 영향을 면밀히 파악하는 것이 필요하며, 경우에 따라 인위적 관리를 통해 복합적 식생구조와 생물다양성 증진 도모를 위한 전략을 검토할 필요가 있다.
연구의 한계
본 연구의 표본지 조사는 제4기 공원자원조사 매뉴얼에서 정한 위치와 개수가 정해져 있는 정점을 중심으로 수행되었다는 한계가 있다. 서언에서 언급했듯이 당해 정점에서 식생뿐만 아니라 식물상, 포유류, 조류, 파충류, 곤충류 조사가 동일한 공간적 범위에서 수행되기 위함이며, 이를 통해 각 정점별 서식지 종합평가가 이뤄진다. 이러한 방법은 동일 서식지 내 다양한 분류군 조사결과를 바탕으로 종합적인 분석‧평가가 이뤄질 수 있다는 장점이 있지만, 식생조사 측면에서는 표본지의 입지‧환경 및 군락 다양성을 고려하기 어렵다는 단점이 있다. 현장에서 식생 여건을 고려하여 조사구의 위치 및 개수를 정하는 방법이 아니기 때문에 소규모의 특수군락, 능선군락, 임연부군락 등이 조사 대상에서 제외될 수 있는 가능성이 있다. 다만, 제4기 공원자원조사의 목적을 고려해보면, 공통된 정점에서 다양한 분류군별 조사가 이뤄졌지만, 당해 지점의 환경요인 정보를 포함하고 있는 분야는 식생 분야가 유일하므로 각 정점별 서식지 종합분석 시 중요한 기초자료로 활용될 수 있을 것으로 판단된다.
또한 지리산국립공원 제3기 공원자원조사의 식생조사 결과와 쌍대비교하기 어렵다는 한계가 있다. 2011년 수행된 제3기 조사는 지리산국립공원 전역에 걸쳐 식분이 균질한 130개의 방형구를 조사하는 등 제4기 조사와는 조사목적, 매목조사 방법, 방형구의 선정기준, 공간적 분포밀도, 입지여건 등 조사체계에 차이가 있다. 제3기 조사는 현존식생도의 작성과 그에 따른 대표군락의 발굴 및 분류가 목적이었으며, 제4기 조사는 대표 유역권별 다양한 분류군별 합동조사를 통해 서식지 종합평가가 목적이었기 때문이다. 따라서 제3기 조사결과를 대조군으로서 비교하는 것은 정보의 왜곡과 곡해를 야기할 수 있다. 동 국립공원의 식생군락의 유의미한 변화상 파악 등 장기 모니터링을 위해 이후 조사에서도 동일 표본지 내 조사가 후속되어야 할 것이다.
적 요
본 연구는 지리산국립공원 전체를 대상으로 표본지 매목조사를 통한 식생군락구조를 파악하고자 하였으며, 지리산국립공원 산림생태계 보전・관리 계획 수립을 위한 최신 자료를 제공하는 목적으로 수행되었다. 종수-면적 곡선을 작성한 결과, 총 40개의 조사구(총 면적 16,000 ㎡)가 지리산국립공원의 출현 종 및 식생현황을 파악하는데 적절한 면적 및 개수였던 것으로 판단된다. TWINSPAN Classification 결과 4개의 군락으로 분류되었다. Ⅰ군락은 들메-당단풍나무군락(Fraxinus mandshurica - Acer pseudosieboldianum Community), Ⅱ군락은 졸참-굴참나무군락(Quercus serrata - Quercus variabilis Community), Ⅲ군락은 신갈나무군락(Quercus mongolica Community), Ⅳ군락은 소나무군락(Pinus densiflora Community)으로 분류되었다. Ⅰ군락은 들메나무군락으로 교목층에 들메나무가 우점하고 있으며, 층층나무와 신갈나무가 경쟁하며 세력을 유지하고 있다. Ⅱ군락은 졸참-굴참나무군락으로 교목층에 굴참나무와 졸참나무가 경쟁하며 우점하고 있으며, 아교목층에서 쪽동백나무와 때죽나무가 경쟁하며 우점하고 있다. Ⅲ군락은 신갈나무군락으로 신갈나무가 교목층에서 67%, 아교목층에서 27%로 높은 우점도를 보이며, 당단풍나무가 아교목층에서 신갈나무와 경쟁하고 있다. Ⅳ군락은 소나무군락으로 교목층에서 소나무가 74%의 높은 우점도를 보이고 있다. 아교목층은 신갈나무가 우점하고 있으며, 개서어나무와 쇠물푸레가 경쟁하며 동반발달하고 있다.
