

서 언
재료 및 방법
실험재료
종자 소독 및 발아실험
종자의 내・외부 형태 관찰
수분 흡수 특성
저온층적처리
고온층적처리
GA3 처리
통계처리
결 과
종자의 내・외부 형태 관찰
수분 흡수 특성
저온층적처리
고온층적처리
GA3 처리
고 찰
적 요
서 언
헐떡이풀속(Tiarella) 식물은 범의귀과(Saxifragaceae)에 속하는 다년생 초본으로 한국, 일본, 중국 및 북미 지역에 9종이 분포하는 것으로 보고되었다(Lee, 2008; The Plant List, 2013; USDA NRCS, 2017). 한반도에 자생하는 Tiarella속 식물로는 헐떡이풀(Tiarellapolyphylla D. Don)이 유일한 종으로 경상북도 울릉도에서만 서식하고 있다. 헐떡이풀은 전통적으로 천식약풀이라고 불리기도 하며 한방에서는 황수지(黃水指)라는 약재로 천식치료제, 염증, 타박상 등의 치료에 이용되었다(NKISBS, 2016). 최근 tiarellic acid라는 성분이 이러한 약리작용을 하는 것으로 밝혀졌으며, 그 효과와 활용에 대해 꾸준히 연구되고 있다(Lee et al., 2012; Moon et al., 2004; Park et al., 1999, 2002). 그러나 제한된 서식지와 개체수의 부족, 그리고 남획의 위협 등으로 산림청에서는 헐떡이풀을 약관심종(Least Concerned, LC)으로 지정하였다(Lee, 2008).
헐떡이풀은 5월 중순부터 무한화서로 흰 꽃을 피우는데, 관상가치가 있어 분화용으로 개발할 수 있고 내음성이 강하여 지피 조경용으로도 개발 가능성이 높다. 이미 외국에서 헐떡이풀속 식물은 다양한 품종으로 개발되어 원예식물로 유통되고 있다(RHS Horticultural Database, 2016). 자생 헐떡이풀에 관련된 증식 및 재배에 관한 연구는 거의 이루어지지 않았다. 북미에 자생하는 헐떡이풀속 식물에 대한 연구는 몇 가지 보고되었다. Kitto and Hoopes (1992)는 T. cordifolia L.의 잎의 절편체 재분화를 통한 기내 미세증식과 삽수의 길이에 따른 기외삽목에 관한 연구를 수행한 바 있다. 종자 휴면에 대해서는 Baskin and Baskin (1988, 2014)이 T. cordifolia L.는 휴면타파를 위해 저온처리가 필요하고 광조건에서 발아가 촉진된다고 보고하였다. Taylor (2001)는 T. trifoliata L.의 발아율이 다른 범의귀과 식물보다 현저히 낮으며, 발아를 위해 저온처리가 필요하다고 보고하였다.
다양한 식물군에서 관찰되는 종자휴면은 휴면유형과 발아 습성에 따라 발아시기를 촉진하거나 지연시키면서 유묘의 생존율에 크게 영향을 미친다(Baskin and Baskin, 2014; Nikolaeva, 1999). 종자휴면은 Baskin and Baskin (2014)에 의해 생리적 휴면(physiological dormancy, PD), 형태적 휴면(morphological dormancy, MD), 형태생리적 휴면(morphophysiological dormancy, MPD), 물리적 휴면(physical dormancy, PY), 그리고 조합휴면(combinational dormancy, PY+PD)으로 구분되었다. 온대지방에서 자생하는 많은 숙근성 식물의 종자는 배가 완전히 자라지 않은 상태로 모식물체에서 탈리되는 것으로 알려져 있다(Martin, 1946). 배의 미성숙으로 인한 휴면은 크게 두 가지로 나눌 수 있다. 발아에 적합한 환경조건에서 배의 발달과 유근의 돌출이 30일 이내에 이루어지는 형태적 휴면(MD)과 발아하기까지 30일에서 수개월 이상의 기간이 소요되고 미숙배가 완전히 자란 후에도 저온 또는 고온층적처리와 같은 별도의 처리를 필요로 하는 형태생리적 휴면(MPD)이 있다(Baskin and Baskin, 2014; Walck et al., 1999).
Baskin and Baskin (2014)은 Tiarella속 종자가 완전히 발달된 배(fully developed embryo)를 가지고 있고 물리적 휴면(PY)은 없으나 고온이나 저온처리를 통해서 휴면이 타파되는 생리적 휴면(PD)을 가진다고 보고하였다. 온대 낙엽수림에 자생하는 범의귀과의 몇 종에 대한 종자휴면 유형이 분류되었다. Heuchera속에서는 H. parviflora와 H. richardsonii 두 종 모두 배가 성숙한 상태이지만 추가적으로 저온처리가 필요한 생리적 휴면(PD)이라고 하였고, Mitella속의 M. diphylla와 Saxifraga속의 S.aizoides 등 11종 또한 생리적 휴면(PD)이라고 보고하였다.
헐떡이풀은 하나의 화서에 15~20개의 소화가 달리고 한 개의 소화에서 약 20여립의 종자를 결실하는데 이러한 개화특성은 실생번식에 유리하다고 할 수 있다. 따라서 본 연구는 헐떡이풀 종자의 휴면타파와 발아특성을 구명하여 대량증식 체계를 확립하고자 수행하였다. 또한 자생 헐떡이풀 종자의 휴면유형을 분류하고 Tiarella속 식물간의 휴면유형 차이를 비교하여 생태적인 연구를 위한 기초자료를 제공하고자 하였다.
재료 및 방법
실험재료
실험에서 사용된 헐떡이풀 종자는 경북 울릉도에서 수집하여 경기도 양평소재의 국립수목원 유용식물증식센터에서 현지외 보존하고 있는 모체에서 2015년 6월 8일 채종하였다. 채종한 종자는 실험실로 가져와서 7~20일 동안 상온건조한 후에 정선하여 실험에 사용하였다.
종자 소독 및 발아실험
정선한 종자는 50 ㎖ 튜브에 넣어 1000 ㎎/L의 베노밀 수화제(Benomyl, FarmHannong, Seoul, Korea)에 8-12시간 동안 침지하였다. 이후 무균작업대에서 75% 에탄올에 1분, 2% NaOCl에 15분 동안 순차적으로 침지하여 표면을 소독하였다. 멸균한 증류수로 5회 이상 수세한 후에 0.7%의 plant agar (Duchefa, Haarlem, Netherlands)만 함유된 무당배지를 25~30 ㎖씩 90 × 15 ㎜ Petri dish에 분주하고 치상한 후, parafilm으로 밀봉하였다. 파종한 종자는 온도 23℃, 습도 50% 조건의 항온 항습이 유지되는 배양실에서 배양하였으며, 40 μ㏖․m-2․s-1 PPFD의 명조건 16시간, 암조건 8시간의 광주기에서 발아를 관찰하였다. 발아는 유근이 1 ㎜ 이상 자란 것으로 정의하였고, 7일 간격으로 발아율을 조사하였다. 최종 발아율(G), 평균발아일수(Mean germination time, MGT), 발아균일도(Germination uniformity, GU), 발아세(Germination speed, GS)를 아래의 식을 이용하여 산출하였다.
• G = (N / S) × 100
• MGT = Σ (TxㆍNx) / N
• GU = Σ [(MGT − Tx)2·Nx] / N – 1
• GS = Σ (Nx / Tx)
(N=총 발아수, S=총 공시 종자수, Nx= 조사 당일의 발아수, Tx= 치상 후 조사일수)
종자의 내・외부 형태 관찰
채종한 종자는 실험실에서 외부형태와 종자를 반으로 자른 내부형태를 USB현미경(AM 3111 Dino-Lite premier, AnMo Electronics Co., Taiwan)을 이용하여 55∼60배율로 관찰하고 촬영하였다. 본 연구에서는 헐떡이풀 종자의 배가 미숙배인지 여부를 판단하기 위해 채종 직후부터 유근이 돌출되기 직전까지 배와 종자의 길이를 측정하였다. 미숙배의 신장을 명확하게 표현하기 위해 배(embryo)와 종자(seed)의 비율인 E:S ratio (Vandelook et al., 2007)를 계산하였다.
수분 흡수 특성
본 실험은 헐떡이풀 종자가 물리적 휴면을 가지고 있는지 판단하기 위해 진행하였다. 정선한 종자들은 1000립 3반복으로 1.5 ㎖ micro 튜브에 넣고 증류수에 침지시켜 수분 흡수 전의 초기 무게와 흡수 후 1, 2, 3, 6, 9, 12, 24, 48시간이 경과되었을 때 무게를 측정하였다. 수분 흡수 정도는 아래의 식을 이용하여 산출하였다.
• %Ws=[(Wi-Wd)/Wd] ×100
(Ws=증가된 종자 무게, Wi=수분 흡수 후 경과 시간 때의 종자 무게, Wd=초기 무게)
저온층적처리
저온층적처리가 휴면타파와 발아에 미치는 영향을 알아보고자 종자를 30립씩 3반복으로 영양분이 무첨가된 배지에 치상하여 5℃에서 0, 12주 저온층적처리를 하였다. 저온층적처리 후 배양실에 배치하여 28주 동안 발아율을 조사하였다.
고온층적처리
고온층적처리가 휴면타파와 발아에 미치는 영향을 알아보고자 종자를 30립씩 3반복으로 무당배지에 치상하여 23℃에서 각각 0, 4, 8, 12주 고온층적처리를 한 후에 5℃에서 8주 동안 저온처리를 하였다. 처리한 종자는 23℃에서 배양하며 16주 동안 발아율을 조사하였다.
GA3 처리
GA3가 휴면타파와 발아에 미치는 영향을 알아보고자 종자를 10, 100, 1000 ㎎/L의 GA3 용액에 침지하여 상온에서 24시간 동안 처리하였다. 증류수에 침지한 것을 대조구(0 ㎎/L)로 두었다. 처리한 종자는 다시 표면소독을 한 후 무당배지에 파종하고 23℃의 배양실에서 16주 동안 발아율을 조사하였다.
통계처리
수분흡수율, 층적처리 그리고 GA3처리에 대하여 조사된 데이터는 SAS 9.4 (SAS Institute Inc., Cary, USA)을 이용하여 분산분석을 하였고, 처리간에 통계적인 유의성은 5% 유의수준에서 Turkey 다중검정을 하였다. 그래프는 Sigma plot 10.0 (SPSS, Inc., Chicago, USA)을 이용하여 작성하였다.
결 과
종자의 내・외부 형태 관찰
종자는 열매(capsule, 삭과)가 주머니 모양으로 싸고 있으며, 종자의 크기는 1 ㎜ 내외로 미립에 속한다(Fig. 1C and Fig. 2A). 종피는 검은색으로 광택이 나며 타원형이다. 채종 직후에 종자의 길이에 대한 배의 크기를 명확하게 표현하기 위해 배(embryo)와 종자(seed)의 비율인 E:S ratio (Vandelook et al., 2007)를 측정하였다. E:S ratio는 0.4 내외로서 배의 크기는 종자 크기의 50%를 넘지 않았고(Fig. 2B), 이를 배양하였을 때 30일이 넘도록 발아하지 않았다(data not shown).


수분 흡수 특성
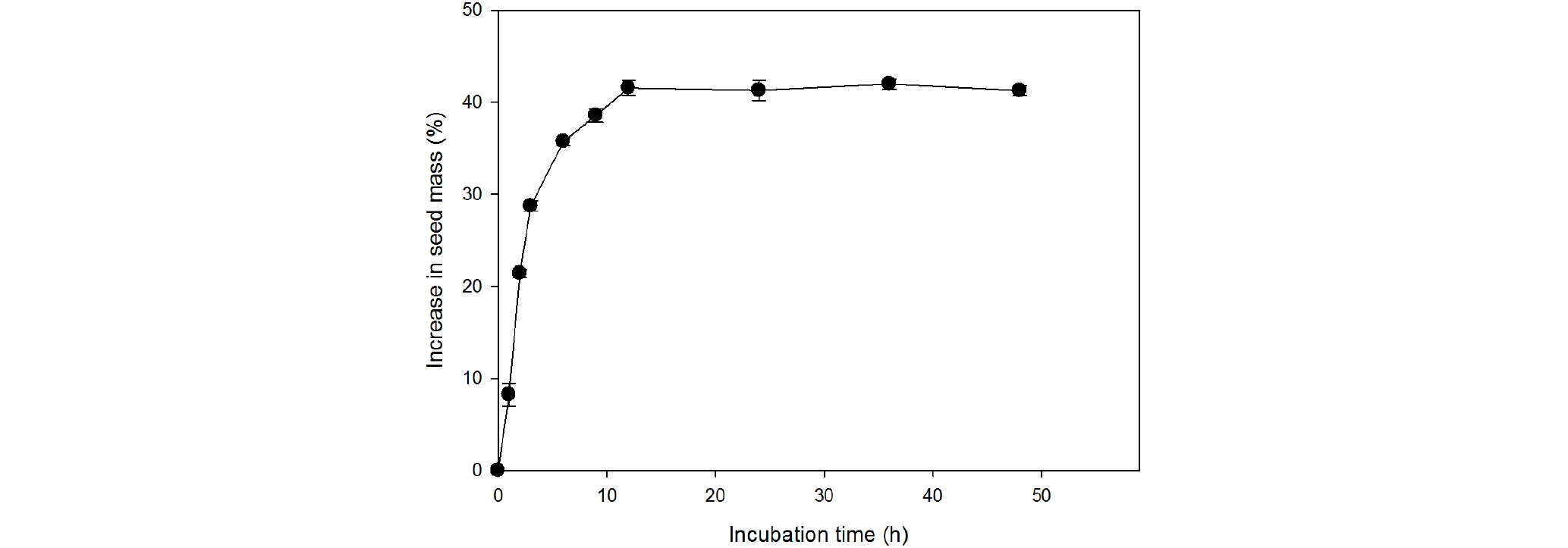
헐떡이풀 종자의 무게는 상온에서 수분 흡수 12시간만에 건조 종자 대비 약 41% 증가하였고 12시간 후부터 포화상태가 되어 더 이상 증가하지 않았다(Fig. 3).
저온층적처리
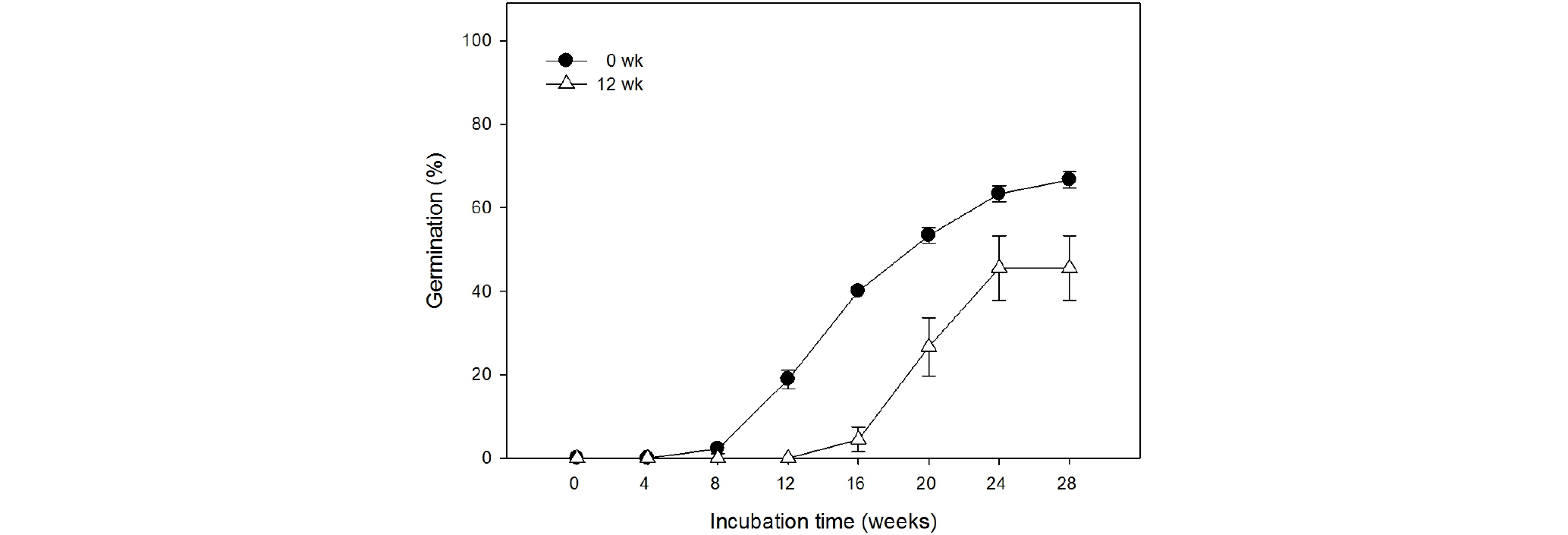
저온층적처리를 하면 무처리구에 비해 발아가 다소 지연되는 것을 확인할 수 있었고 평균발아일수가 무처리구는 111.3일이었으며 12주 저온처리구는 135.1일로 나타났다(Table 1, Fig. 4). 무처리구가 배양 8주차 부터 발아를 시작한 반면, 12주 저온처리구에서는 배양 16주 후부터 발아가 시작되었고, 최종 발아율도 무처리구 66.7%에 비해 45.6%로 낮게 나타났다. 발아 과정에서 종자는 부피가 커지고 종피가 찢어진 후에 유근이 돌출하면서 발아가 이루어졌다(Fig. 2C-D). 이때 종자를 잘라 배를 관찰하였을 때, 배의 길이가 신장한 것을 확인할 수 있었다. E:S ratio는 채종 직후 0.4 내외에서 발아 직전 0.7 내외로 약 75% 증가하였다.
Table 1. Effects of cold stratification on germination characteristics of Tiarella polyphylla D.Don seeds
| Cold stratification Periods (weeks) | Final germination (%) | Mean germination time (day) | Germination uniformity | Germination speed |
| 0 | 66.7 | 111.3 | 742.4 | 0.197 |
| 12 | 45.6 | 135.1 | 88.5 | 0.113 |
| Significance | NSz | **z | ** | ** |
고온층적처리
고온층적 처리기간이 길수록 발아가 빨리 시작되고 최종 발아율이 높은 경향을 보였다(Fig. 5). 무처리구가 배양 12주 후부터 발아가 시작되는 반면, 4주 고온처리구는 8주만에, 8주와 12주 고온처리구는 4주만에 발아를 시작하였다. 발아율도 고온층적 처리기간이 길수록 높게 나타났다. 배양 16주 후에 측정한 발아율은 12주 처리구가 51.1%로 가장 높았으며, 8주 처리구는 20.0%, 4주처리구는 24.4%, 무처리구는 13.3%로 나타났다.
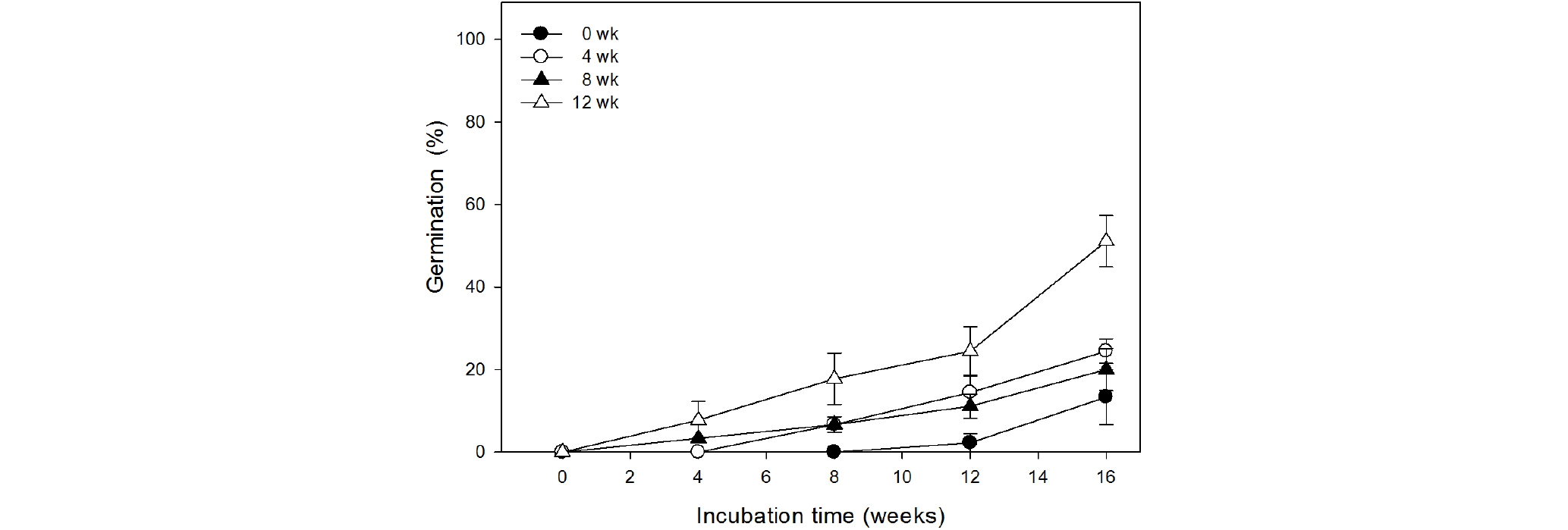
Fig. 5.
Percentage germination of Tiarella polyphylla D.Don seeds as affected by difference warm stratification period. The seeds were warm stratified at 23℃ for 0, 4, 8, or 12 weeks. After warm stratification for each period, all seeds were cold stratified at 5℃ for 8 weeks and then moved to 23℃ for incubation. Vertical bars represent SE (n=3).
GA3 처리
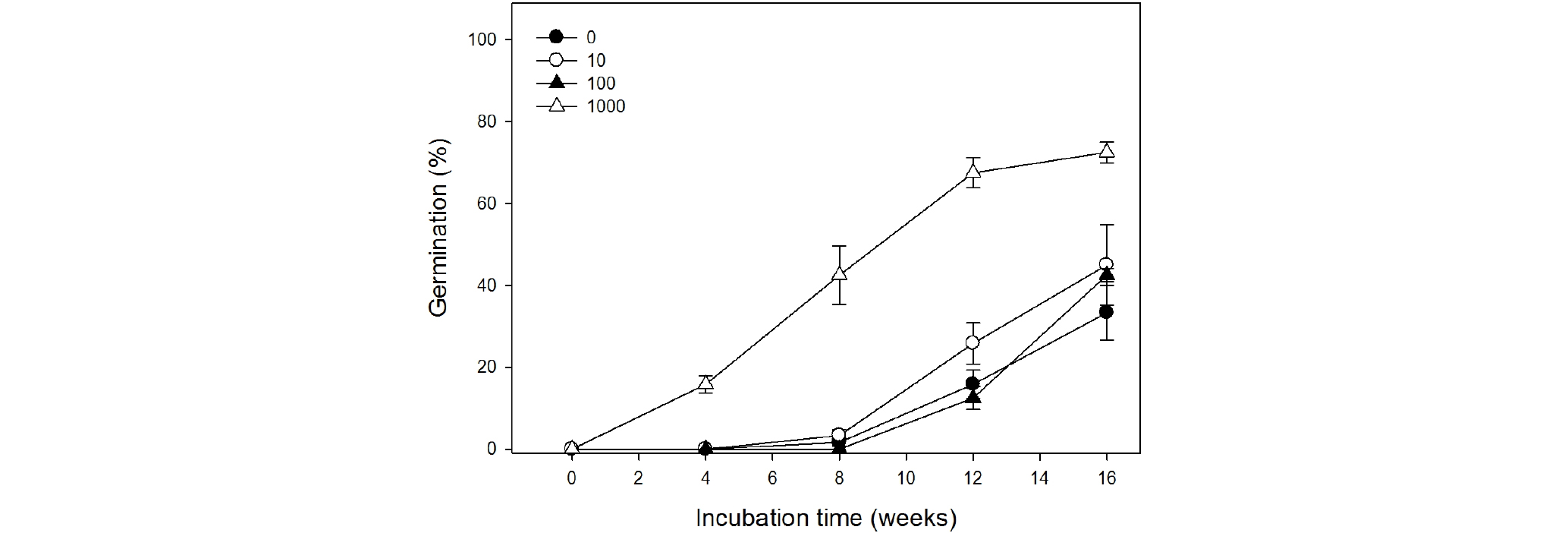
GA3 0, 10, 100 ㎎/L 농도처리의 최종 발아율은 33.3, 45.0, 42.5%였으나 GA3 1000 ㎎/L 처리구에서는 최종 발아율이 72.5%로 가장 높게 나타났다(Table 2, Fig. 6). 또한 GA3 1000 ㎎/L 처리구에서 평균발아일수(MGT)가 53.7일로 가장 짧았으며, 발아균일도(GU)도 388.7로 가장 균일하게 나타났다. GA3 0, 10, 100 ㎎/L 농도처리의 평균발아일수는 89.5, 81.5, 95.4일로 GA3 1000 ㎎/L 처리구에 비해 길게 나타났다.
Table 2. Effects of GA3 treatment on germination characteristics of Tiarella polyphylla D.Don seeds
고 찰
Baskin and Baskin (2014)은 종자휴면의 유형을 생리적 휴면(PD), 형태적 휴면(MD), 형태생리적 휴면(MPD), 물리적 휴면(PY), 그리고 조합휴면(PY+PD) 등 크게 다섯 가지로 분류하고 각각의 유형 내에서도 휴면의 깊이에 따라 여러 가지 유형으로 세분화하였다. 물리적으로 종자내부로 수분이 흡수되지 못하여 발아가 지연되는 휴면을 물리적 휴면(PY)이라 하며, 종자의 초기 무게에 비해 수분을 흡수 후 종자의 무게가 약 20% 이상 증가하면 투수성이 있다고 판단한다(Baskin and Baskin, 2004). 헐떡이풀 종자는 두 시간 만에 초기 무게에 비해 증가하였고, 12시간 경과 후 약 41% 증가하여 포화상태에 도달하였다(Fig. 3). 따라서 헐떡이풀 종자는 물리적 휴면이 없는 것으로 판단된다. 미세종자에 속한 자생식물인 난쟁이바위솔(Cho et al., 2014), 숙은노루오줌(Jang et al., 2016) 등에서도 침지시간에 따라 종자의 무게가 증가함으로써 물리적 휴면과 복합휴면이 없다고 보고하였다.
Martin (1946)은 종자 내부의 배 형태를 크게 기저형(basal), 지협형(peripheral), 그리고 중축형(axile)의 세 가지로 분류하였다. 기저형은 일반적으로 배가 작고 종자의 하단부에 위치하고 크기가 종자의 50%를 넘지 않으며, 그 중 작은 타원형 배가 종자의 하부에 위치한 rudimentary 타입이 대표적인 형태이다. 지협형은 배가 길고 종피를 따라 휘거나 말려있는 형태이다. 중축형은 배가 작거나 중앙부에 길게 위치하거나 또는 휘어있는 등 다양한 형태가 있는데, 그 중 작고 길쭉한 형태의 배가 종자의 중앙부에 위치한 linear 타입을 대표적인 형태로 보고하였다. 헐떡이풀과 동속 식물인 Tiarella cordifolia L.의 배 형태는 범의귀과의 Chrysospleniumamericanum와 Hydrangeaarborescens와 더불어 중축형(axile)의 dwarf 타입으로 분류하였다(Baskin and Baskin, 2007; Martin, 1946). 자생 헐떡이풀 종자의 배는 종자의 중앙에서 약간 길쭉한 모양으로 하단부에 위치하고 있어(Fig. 2B) 중축형(axile)의 linear 타입으로 판단된다.
온대지역에 자생하는 많은 초본성 식물들의 종자는 배가 충분하게 자라지 못한 상태로 모체에서 떨어지는 것으로 보인다(Baskin and Baskin, 1988). 이러한 미숙배(underdeveloped embryo)를 가지는 종자는 형태적 휴면(MD) 또는 형태생리적 휴면(MPD)을 가지고 있다. 헐떡이풀 종자는 채종 직후에 배의 길이를 확인하였을 때, 종자의 약 40% 크기였으나 발아 직전에 약 70%의 크기로 배가 신장하여 채종 시기의 헐떡이풀 종자는 미숙배임을 판단할 수 있다(Fig. 2B and 2D). 또한 헐떡이풀 종자는 파종하고 30일이 경과한 후에도 배의 신장과 발아가 이루어지지 않았기 때문에 생리적 휴면이 있다고 판단할 수 있다. 따라서 미숙배를 가진 헐떡이풀 종자는 형태생리적 휴면(MPD)으로 분류할 수 있다. 그러나 동속 식물인 Tiarella cordifolia L.와 Tiarella trifoliata L. 종자는 저온처리를 통해 휴면이 타파되는 생리적 휴면(PD)을 가진다고 보고된 바 있어 본 연구 결과와는 상반된 결과임을 알 수 있다(Baskin and Baskin, 1988, 2014; Taylor, 2001). Baskin and Baskin (1988)과 Taylor (2001)는 각각의 헐떡이풀속 식물 종자의 배 길이나 발달에 대한 구체적인 데이터를 제시하지 않았다. 동속 식물 내에서 휴면유형이 다른 경우는 종종 보고되었다. 진달래과(Ericaceae)의 Leucopogonconostephioides, L. polymorphus는 생리적 휴면(PD)으로 분류하였지만(Kullmann, 1981), L. esquamatus, L. exolasius, L. setiger는 미숙배를 가진 형태생리적 휴면(MPD)이라고 분류하였다(Ooi et al., 2006). 또 물푸레나무과(Oleaceae) 물푸레나무속(Fraxinus) 식물 내에서도 F. Americana (Steinbauer, 1937)와 F. angustifolia (Piotto, 1994) 등은 생리적 휴면(PD)으로 분류하였지만, F. excelsior (Villiers and Wareing, 1964), F. nigra (Steinbauer, 1937; Benedict and David, 2003)는 형태생리적 휴면(MPD)으로 분류하였다.
형태생리적 휴면은 휴면타파를 위한 온도와 배가 발달하는데 필요한 온도, 그리고 GA (gibberellic acid)가 휴면을 타파하는지에 따라서 여덟 가지로 세분화된다(Baskin and Baskin, 2014). 배가 발달하는 온도가 상대적 고온(≥15℃)인 경우엔 simple-type MPD로, 상대적 저온(≤10℃)인 경우 complex- type MPD로 나눌 수 있으며, 각각의 유형 내에서 생리적 휴면의 깊이에 따라 non-deep MPD, intermediate MPD, deep MPD로 분류된다. 헐떡이풀 종자의 경우 12주 동안 저온층적처리를 하였을 때 오히려 발아가 지연되고 발아율이 무처리구에 비해 낮아졌다(Table 1 and Fig. 4). 그러나 저온층적처리를 하지 않더라도 23℃에 배양하였을 때 발아율이 점차적으로 증가하였다(Fig. 4). 이는 23℃의 상대적 고온에서 지속적으로 배가 자란다는 것을 의미한다. 이를 통해 헐떡이풀 종자의 배 발달을 위해서 요구되는 온도가 고온인 simple-type MPD로 판단할 수 있다. GA는 종자의 휴면타파와 배의 발달, 그리고 발아를 조절하는 호르몬이다(Baskin and Baskin, 2014; Finch-Savage, 2006). 헐떡이풀 종자의 생리적 휴면 깊이를 알아보고 배 발달과 휴면타파에 필요한 고온층적처리를 대체할 수 있는지 확인하고자 GA3를 농도 별로 처리하였을 때, 최종 발아율이 무처리구는 33.3%, GA3 1000 ㎎/L 처리구는 72.5%로 나타났다. 평균발아일수도 89.5일(무처리구)에서 53.7일(GA3 1000)로 약 2배 촉진되었다(Table 2 and Fig. 6). 따라서 헐떡이풀 종자는 미숙배가 상대적 고온에서 자라고 GA3로 휴면이 타파되는 non-deep simple MPD로 분류할 수 있다.
헐떡이풀은 GA3처리를 통해 종자의 휴면을 효과적으로 타파할 수 있었으며, 이러한 결과를 통해 헐떡이풀의 대량증식 체계를 구축하기 위한 기초자료를 확보하였다. 더 나아가 본 연구에서 밝혀진 헐떡이풀의 종자 휴면유형은 형태생리적 휴면(MPD)으로서 기존에 보고된 다른 Tiarella속 식물의 생리적 휴면(PD)과는 다르게 나타났다. 이러한 차이가 지리적 분포에 따른 환경에 기인한 차이인지, 아니면 동일 속 내의 진화과정에 의한 것인지 추가적인 검토가 필요할 것이라 사료된다.
적 요
헐떡이풀은 우리나라에서 울릉도에서만 분포하는 자생식물로 전통적으로 약용으로 쓰여왔으며, 관상식물로의 활용도 기대되는 자원식물이다. 본 연구는 헐떡이풀 종자의 휴면타파와 발아를 위한 조건을 확립하고 종자휴면 유형을 분류하고자 수행되었다. 실험은 헐떡이풀 종자에 저온층적처리(5℃에서 0, 12주)와 고온층적처리(23℃에서 0, 4, 8, 12주 처리 후에 5℃에서 8주, 다시 23℃에서 배양함), GA3 (0, 10, 100, 1000 ㎎/L)처리를 수행하였다. 이렇게 처리한 종자는 무균배지에 소독하여 파종하고 온도 23℃, 광도 40 μ㏖․m-2․s-1 PPFD, 일장 16시간 조건의 배양실에서 배양하였다. 헐떡이풀 종자는 자연상태에서 미숙배인 상태로 탈리되며, 종자 내부로의 수분흡수에 대한 물리적인 장벽은 없었다. 그러나 아무 처리 없이 파종하였을 때 30일이 지나도록 전혀 발아하지 않았다. 따라서 헐떡이풀 종자는 형태적 휴면(MD)과 생리적 휴면(PD)을 가지고 있는 것으로 판단하였다. 저온층적처리(0, 12주) 실험의 최종 발아율은 각각 66.7%, 45.6%로 나타났다. 저온처리는 오히려 발아를 약 3주 정도 지연시켰다. 고온층적처리(0, 4, 8, 12주) 실험에서는 최종 발아율이 각각 13.3%, 24.4%, 20.0%, 51.1%로 나타났다. 헐떡이풀 종자의 배는 상대적 고온에서 발달하였다. GA3 처리는 종자의 휴면을 극복하고 발아를 촉진하였다. GA3 처리에서의 최종 발아율은 0, 10, 100, 1000 ㎎/L 처리구에 대하여 각각 33.3%, 45.0%, 42.5%, 72.5%로 나타났다. 위의 결과를 종합하여 헐떡이풀 종자는 non-deep simple 형태생리적 휴면(MPD)을 가지고 있으며, GA3 처리를 통해 고온을 대체하고 종자의 휴면을 타파할 수 있었다. 이는 한국에 자생하는 헐떡이풀속의 종자휴면에 대해 처음으로 보고한 논문이다.
