

서 론
재료 및 방법
조사지 선정
입지환경 조사
식생조사
식생군집구조
통계 분석
결과 및 고찰
임분개황
토양 이화학적 분석
기상현황
군집구조분석
종다양성
유사도지수
유집분석에 의한 군집구조 분석
상관관계분석
적 요
서 론
개비자나무(Cephalotaxus koreana Nakai)는 개비자나무과(Cephalotaxaceae)에 속하는 상록관목으로 경기, 충북 이남의 계곡이나 숲의 습윤한 곳에 자생하는 우리나라 특산 식물이다. 수고 3~6 m 정도로 자라며, 잎은 선형으로 두 줄로 배열하며, 주맥이 양면에 도드라지고 잎 뒷면에는 두 줄로 된 기공선이 있으며, 잎자루는 없다. 꽃은 암수딴그루로 4월에 개화하며, 수꽃은 편구형으로 연한 갈색이며 10여 개의 포로 싸여 한 꽃 자루에 20~30개씩 달리고, 암꽃은 2개씩 달리며 10개의 뾰족한 녹색포로 싸인다(Korea National Arboretum, 2016). 개비자나무에 대한 연구는 주로 화학적 성질, 항암성분에 대해 외국에서 먼저 밝혀졌다(Powell et al., 1972; Ohnamam and Holland, 1985; Edward, 1999). 암을 극복하고자 하는 인류는 유용자생식물을 이용한 항암제 개발에 초점을 맞추고 있는 가운데 개비자나무에서 추출된 항암물질은 기존 항암제에 비해 물에 대한 용해도가 높다고 보고되어, 단점을 극복할 수 있는 상품화 가능한 차세대 항암제로 주목받고 있다(Jingyi et al., 2000). Park et al. (1996)은 국내 개비자나무에서도 항암활성을 가지는 homoharringtonine과, harringtonine이 있다고 보고하였고, 부위별 항암성분은 개비자나무의 잎 추출물이 줄기, 열매에서 보다 2~3배 정도 많이 함유하고 있다고 보고하여(Kim et al., 2000; Jung et al., 2005) 유용자생식물로서 활용 가치가 대단히 높다.
한편 생물 다양성협약(Convention on Biological Diversity, 1992)의 채택으로 식물자원의 배타적 권리가 인정됨에 따라 국가 간 식물유전자원의 확보와 산업화 경쟁이 심화되고 있으며, 각종 국제협약단체는 식물자원의 이용 및 개발에 대한 상업적 권리를 제도화하고 있다(National Forest Seed Variety Center, 2009). 또한 향후 50년 이내에 지구상에 분포하는 25만종의 관속식물 중 약 20%가 멸종될 것이라고 추정된 바(Falk and Olwell, 1992), 생물다양성을 최대한 유지시킬 수 있는 보전전략 및 생물종의 증식과 자생지의 특성에 관심이 높아지고 있다(Kim, 2007). 최근 우리나라에서도 멸종위기 및 자생식물의 서식지 특성과 보전방향에 대한 연구의 일환으로 차나무 자생지의 분포 특이성(Eom and Kim, 2017), Yoon et al. (2015)의 개병풍 자생지 환경특성, Kim et al. (2014)의 가침박달 자생지 환경특성, Byun et al. (2013)의 애기송이풀 자생지 식생구조 연구를 비롯하여, 특정 지역 산림의 식물상에 관한 연구 등 식물자원 보전을 위한 연구가 진행 중이다(Kim et al., 2018; Sin et al., 2018; Han et al., 2018).
개비자나무는 생장시기 동안 종자결실 주기가 느리고, 자웅이주이며, 서식지가 단편화 되는 등 멸종 위기에 처해 있다고 보고되었다(Fu et al., 1999). 이러한 상황에서 개비자나무에 대한 연구로는 항암성분에 관한 연구에 초점이 맞춰 이루어졌을 뿐 국내 자생하는 개비자나무의 생육환경과 군집구조에 대한 조사는 이루어지지 않았다. 다만 Jung et al. (2009)은 신규 항암소재로서 개비자나무의 대량 생산을 위해 삽목증식 특성에 대해 연구한 결과 IBA 100 ㎎/L 처리구에서 88.5%의 높은 발근율을 보고하였고, 이는 종 보전 측면에서 유리할 것으로 판단된다.
본 연구에서는 개비자나무 자생지의 입지특성, 토양환경, 식생구조 등을 파악하여 개비자나무 자생지의 생육환경을 구명하고 지속적인 보전과 생태환경복원을 위한 기초자료를 제공하고자 실시하였다.
재료 및 방법
조사지 선정
개비자나무의 조사지는 인위적 교란이 없고, 해당 수종이 20본 이상 생육하는 지역으로 충북에 충주 적보산, 괴산 보개산, 경남에 거창 금원산, 전남에 구례 지리산, 광양 백운산, 해남 두륜산으로 6개 집단을 선정하였다(Fig. 1, Table 1).
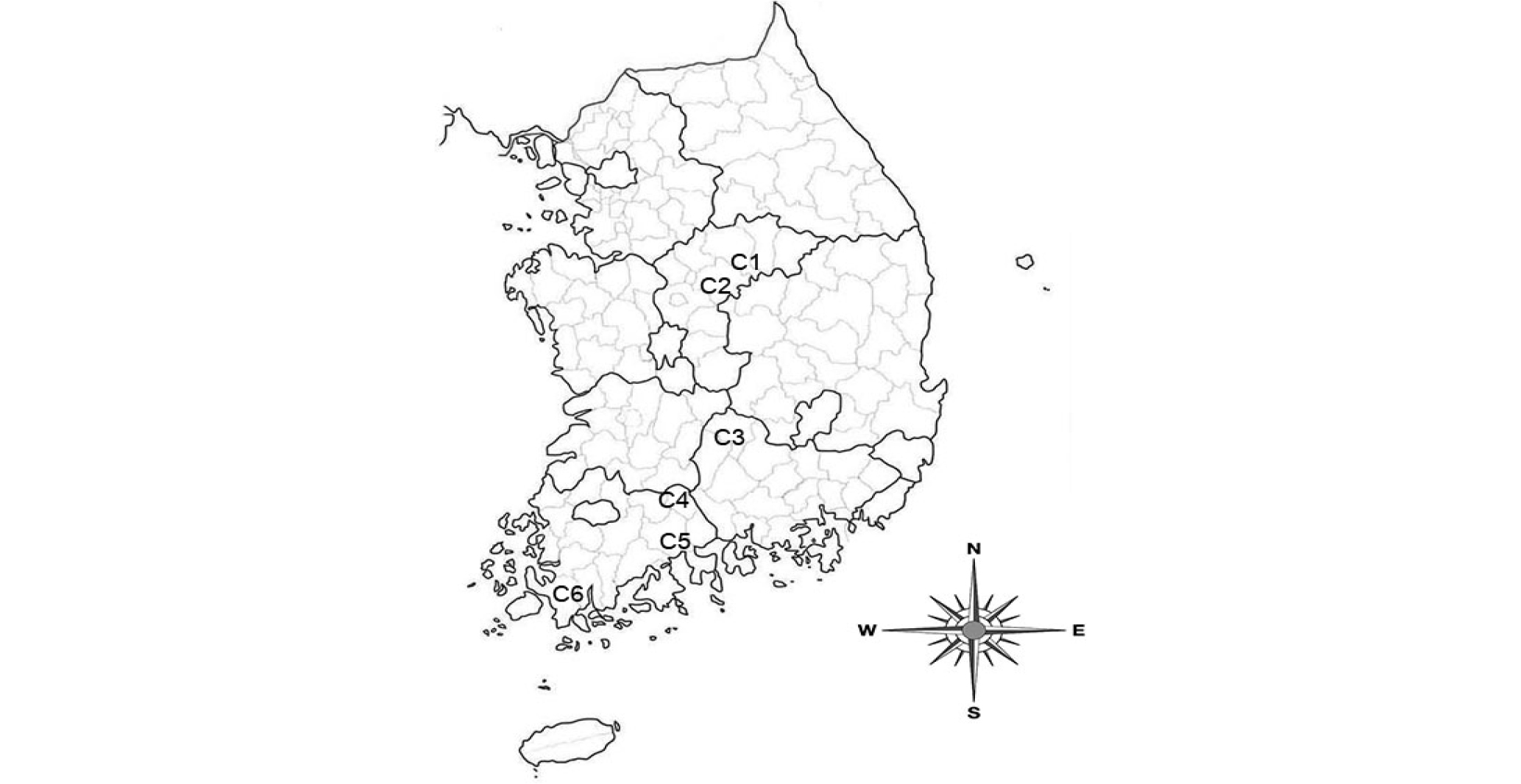
Table 1. Location of Cephalotaxus koreana habitats
입지환경 조사
조사지의 해발고, 경사, 방위를 측정하고, 생육하고 있는 개비자나무의 수고와 직경을 조사하였다. 토양의 화학적 성분을 조사하기 위해 각 조사구 내에서 낙엽층을 제거한 후 토양을 채취하여 산도(pH)는 pH meter법, 유기물함량(O.M)과 전질소(T-N)는 건식산화법, 유효인산(Avail P2O5)은 Lancaster법, 양이온치환용량(CEC)은 1N-초산암모니움 침출법, 치환성 양이온(K⁺, Ca⁺, Mg2⁺)은 원자흡광광도법을 이용하여 분석하였다.
기상자료는 2011년부터 2015년까지 5년간의 조사지의 월 평균기온, 온량지수(Warmth index, WI), 한랭지수(Coldness index, CI), 홀드리즈 지수(Holdridge, 1947), 강수량, 습도 등 수목생육과 관계가 있는 요소들을 기상청(Korea Meteorological Administration, 2016) 자료를 이용하여 산출하였다.
식생조사
선정된 개비자나무 조사지에 각각 10×10 m 방형구 3개씩 총 18개의 조사구를 설치한 후 Braun-Blanquet법(1964)에 의해 교목층(수고 8 m 이상), 아교목층(2~8 m), 관목층(2 m 이하)로 층위를 구분하여 식생조사를 실시하였다.
식생군집구조
각 조사지별 임관층의 종간 상대적 우위를 비교하기 위하여 조사구내 출현하는 모든 수목을 대상으로 상대밀도, 상대피도를 산출하고 이를 바탕으로 임관층위별 중요치로 표현되는 상대중요치(Importance value : IV)를 구하였다(Curtis and McIntosh, 1951; Brower and Zar, 1977). 이후 개체들의 크기를 고려하여 수관층위별로 가중치를 부여한 후 평균상대중요치(Mean importance value : MIV)를 구하여(Park et al., 1987) 유집분석에 이용하였다.
종 구성 상태의 다양한 정도와 중요치를 분석하기 위하여 Shannon-Weaver의 수식(1949)을 이용하여 종다양도(H')와 최대종다양도(H' max)를 산출하였으며(Pielou, 1975), 각 수목에 대한 개체들의 분포를 의미하는 균재도(Evenness, J'), 우점도(Dominance, 1-J')를 계산하였고, 조사지별 종 구성의 유사성을 비교하기 위하여 Whittaker의 방법으로 유사도지수(similarity index)를 구하였다(Whittaker, 1956).
통계 분석
조사된 자료는 SPSS program(Statistical Package for Social Science, ver. 22.)을 이용하여 상관관계를 분석하였고, 조사지별 평균상대중요치 값을 이용하여 유집분석을 실시하였다.
결과 및 고찰
임분개황
각 조사지별 해발고, 경사, 방위, 평균수고, 평균흉고직경, 흉고단면적 및 개비자나무 생육상태 등 임분개황을 조사한 결과는 Table 2와 같다.
Table 2. General description of the Cephalotaxus koreana habitats
해발고도 148~835 m, 경사 12~32°로 다양하게 나타났으며, 방위도 일정한 경향이 발견되지 않았다. 평균수고는 교목층의 경우 9~14 m, 아교목층 4~5 m의 범위를 보였으며, 임지의 생산성을 나타내는 흉고단면적은 교목층에서 30.2~48.0 ㎡/ha, 아교목층 1.34~4.79 ㎡/ha로 나타났다. 특히 보개산과 지리산은 암석이 많이 분포하는 암석지였으며, 금원산과 백운산은 계곡부에 위치하여 토양습도가 높은 특징이 나타났다.
조사지별 생육하고 있는 개비자나무에 대해 직경, 수고를 조사한 결과 적보산, 보개산, 금원산, 백운산 집단에서는 근원직경 2 ㎝, 수고 1.5 m이하로 관목층에 서식하여 비교적 작게 자랐으나 개체수가 많고 집단적으로 분포한 반면에 지리산, 두륜산에서는 흉고직경 6 ㎝ 이상, 수고 3 m로 아교목층에 서식하여 다른 집단보다 크게 자랐으나 개체수가 적고 국부적으로 나타났다. 수고 및 흉고직경에 차이가 나는 것은 남부지역으로 내려갈수록 수령이 오래되고, 중부지역으로 올라갈수록 5년 이하의 개비자나무가 주로 분포한다고 보고한 결과와 마찬가지로 수령에 의한 차이로 판단된다(Jung, 2005). 또한 따뜻한 지역에서 개비자나무의 생육이 왕성하다고 보고한 선행연구 결과(Fu et al., 1999)와 마찬가지로 지리산과 두륜산의 경우 남부지역으로 따뜻한 기후에 속하면서 남향사면의 방위를 나타내어 온도 등 수목의 생육에 유리한 환경을 제공하였기 때문으로 판단된다.
토양 이화학적 분석
산림군락의 분포를 결정하는 요인 중의 하나로 개비자나무 6개 조사지에 대한 산림토양의 이화학적 분석을 실시한 결과는 Table 3과 같다.
Table 3. Soil characteristics of Cephalotaxus koreana habitats
토양 pH는 4.7~5.9까지 분포하여 일반산림 토양의 평균 pH인 5.5에 비해 낮은 수치를 나타냈으며, 이는 개비자나무가 산악지역 계곡부와 경사지에 주로 분포하여 염기성양이온 용탈이 심하기 때문으로 판단된다. 유기물함량은 11.1%로 높은 수치를 나타냈으며 자연림으로서 상층 및 중층으로부터 낙엽활엽수의 낙엽낙지와 지하부 세근 등에 의한 유기물 유입이 활발하게 발생하기 때문으로 판단된다. 한편 보개산과 지리산의 경우 5.95%, 5.72%를 나타내어 비교적 적은 유기물 함량을 나타냈다. 유기물함량은 생산량과 유기물분해에 의해 기인한 것으로(Nakane et al., 1986) 임내에서의 온도와 수분의 증가는 유기물 층의 분해를 촉진한다고 하였는데(Hwang et al., 2006), 보개산과 지리산은 서식지 내 암석의 비율이 높아 유기물의 축적량이 적고, 유기물 분해가 느리기 때문인 것으로 판단된다. 양이온치환용량의 경우 평균 17.4 cmolc/㎏-1으로 우리나라 산림토양값 12.5 cmolc/㎏-1 보다 높게 나타났으며, 전질소(T.N.)의 경우 평균 0.48%로 우리나라 산림토양 평균 0.19%에 비해 높았다(Jeong et al., 2002). K+, Ca2+, Mg2+ 등 치환성양이온 함량은 각각 0.36 cmolc/㎏, 3.36 cmolc/㎏, 0.81 cmolc/㎏으로 우리나라 산림평균보다 높게 나타났다. pH를 제외한 대부분의 토양 특성이 산림 평균보다 높게 나타난 것은 개비자나무가 낙엽의 생산량이 많은 낙엽활엽수림에 자생하고 유기물 축적이 이루어지는 계곡부와 산복사면에 주로 서식하기 때문인 것으로 판단된다.
기상현황
조사지의 기상자료는 2011년~2015년까지 5년간 각 조사지의 인접한 기상대에서 측정된 기상자료를 활용하였다. 즉 적보산은 충주, 보개산은 괴산, 금원산은 거창, 지리산은 남원, 백운산은 광양, 두륜산은 해남의 기상대에서 측정한 자료를 활용하여 온량지수, 한랭지수, Holdridge지수, 강수량, 습도 등을 산출하였다(Table 4).
Table 4. Climatic indicators using adjacent network in the Cephalotaxus koreana habitats from 2011~2015
조사지의 연평균기온은 9.7~12.4℃ 범위를 나타냈고, 연평균강수량은 1,220~1,591 ㎜, Holdridge지수는 10.0~12.5℃, 온량지수는 81.0~101.4℃, 한랭지수 -28.3~-13.8, 연평균 상대습도는 65.3~74.5로 나타났다(Table 3). Yim (1977)은 우리나라의 식생대를 온량지수와 한랭지수에 의하여 구분하고 온량지수에 따른 식생연속변화에 의하여 온량지수 30~70℃의 범위는 아고산대림, 50~90℃는 냉온대림, 80~100℃는 온난대 낙엽활엽수림, 100~120℃는 온난대 상록활엽수림의 4개 식생대로 구분하였다. 이상의 결과를 종합하면 조사된 개비자나무 집단을 온량지수 및 한랭지수 등에 의해 구분할 때 온난대 낙엽활엽수림에 속하는 것으로 나타났다.
군집구조분석
개비자나무 6개 조사집단에 대한 식생구조를 분석하기 위하여 출현종수에 의한 식생층위별 중요치(importance value : IV)와 평균상대중요치(Mean of importance value : MIV)를 산출한 결과 목본은 교목층에서 관목층까지 총 52종이 나타났다(Appendix 1, Appendix 2).
개비자나무 집단 중 충북 충주 적보산의 교목층 중요치는 떡갈나무가 65%, 쪽동백나무 17.1% 순으로 우점하면서 총 5종이 출현하였고, 아교목층에서는 비목나무 45.2%, 노린재나무 22.4% 등 4종이었으며, 관목층에서는 개비자나무 54.7%, 비목나무 15.3% 등 총 10종이 출현하였다.
충북 괴산군 보개산 집단의 교목층에서는 고로쇠나무 30.1%, 느티나무 21.9% 순으로 총 7종이 출현하였고, 아교목층에는 비목나무 40.8%, 물푸레나무 20.8% 등 8종이었으며 관목층은 개비자나무가 68.9%로 가장 높은 가운데 병꽃나무 8.0% 등 총 10종이 나타났다.
경남 거창 금원산 조사지의 교목층 중요치는 물푸레나무 47.0%, 당단풍나무 22.5% 등 총 8종이 출현하였으며, 아교목층은 당단풍나무 51.1%, 쪽동백나무 19.6% 등 총 6종이었으며, 관목층의 경우 개비자나무가 73.0%로 가장 높은 가운데 총 6종이 출현하였다. 금원산의 경우 개비자나무의 평균 흉고직경 2.25 ㎝, 수고 1.39 m로 관목층에 분포하면서 근맹아를 통한 번식이 활발하여 많은 개체수가 관찰되었으며, 조사지 중 관목층에서 개비자나무의 중요치가 가장 높게 나타났다.
전남 구례군 지리산 조사지의 교목층 중요치는 졸참나무 17.0%, 잔털벚나무 14.7%, 때죽나무 14.4% 등 11종이 균등하게 출현하였고, 아교목층은 개비자나무 49.5%, 사람주나무 16.4% 등 10종이었으며, 관목층은 산초나무 17.3%, 비목나무 12.7% 등 총 13종이 출현하였다.
광양 백운산의 조사지의 교목층에서는 느티나무가 39.9%, 갈참나무 26.1% 등으로 총 6종이 나타났으며, 아교목층은 당단풍나무27.4%, 사람주나무 26.9% 등 총 7종이 출현하였다. 관목층은 개비자나무 36.5%, 박쥐나무 23.0% 등 총 9종이 나타났다.
해남 두륜산 조사지의 교목층은 상수리나무가 24.7%, 신갈나무 16.9% 순으로 총 10종이 나타났으며, 아교목층은 개비자나무가 32.3%, 때죽나무 18.3% 등 총 6종이 나타났다. 총 8종이 출현한 관목층은 개비자나무가 23.7%, 때죽나무 18.3%, 생달나무 16.3%, 생강나무 12.3% 순으로 나타났다.
개비자나무가 집단을 이루는 조사지 상층수목의 우점종은 적보산에서 떡갈나무, 쪽동백나무, 보개산에서 고로쇠나무, 느티나무, 금원산에서 물푸레나무, 당단풍나무, 지리산에서는 졸참나무, 잔털벚나무, 백운산에서는 느티나무, 갈참나무, 두륜산에서는 상수리나무, 신갈나무가 우점하여 상층식생이 공통적으로 우점하고 있는 수종은 찾아볼 수 없었으며, 낙엽활엽수종 중 참나무류가 우점하고 있었다. 또한 교목층의 종이 많고 비교적 균등하게 우점하고 있는 지리산, 두륜산에서 개비자나무의 생육이 우수한 것으로 나타났으며, 이러한 결과들을 종합해 볼 때 개비자나무는 상층수목에 의해 피음도가 높은 지역의 계곡부나 습윤지에 생육하고 있는 것으로 판단된다.
조사 집단에서 출현한 52종 중에서 6개의 조사지에서 공통으로 출현한 종은 개비자나무와 당단풍나무 2종외에는 없었다. 5개 집단에서 공통으로 출현한 종은 비목나무, 작살나무, 생강나무 3종이었고, 느티나무, 고로쇠나무, 쪽동백나무, 사람주나무는 4개 조사지에서 공통으로 출현하였다.
종다양성
종 구성 상태의 다양한 정도와 중요치를 분석하기 위해 조사 지역별 출현종수와 Shannon의 수식을 적용하여 종다양도(H'), 각 수목에 대한 개체들의 분포정도를 의미하는 균재도(J'), 우점도(D) 및 최대종다양도(H'max)를 조사한 결과는 Table 5와 같다.
Table 5. Values of species diversity in the Cephalotaxus koreana habitats
수관층 전체의 조사지별 출현종수는 15~26종으로 지리산이 26종으로 가장 많게 나타났으며, 적보산이 15종으로 가장 적게 나타났다. 종간경쟁의 정도를 나타내는 종다양도는 0.854~ 1.234의 범위로 두륜산이 1.234, 지리산 1.229 순으로 나타났으며, 백운산이 0.854로 가장 낮게 나타났다. 균재도(J')는 상대적인 종의 다양도를 뜻하는 것으로 1에 가까울수록 종별 개체수가 균일한 상태를 나타내는데(Brower and Zar, 1977) 개비자나무의 균재도는 0.654~0.933의 범위를 나타냈다. 이는 산겨릅나무 자생지(Son et al., 2016)의 균재도 0.871~0.918, 가침박달(Kim et al., 2014) 자생지의 균재도 0.77~0.92에 비해 다소 낮은 값이나 차나무 자생지에서 균재도는 0.509~0.876 이라고 보고한 결과보다 다소 높게 나타났으며(Oh, 2008), 비교적 임분이 여러 종에 의해 균일하게 분포하여 상호 경쟁하고 있는 상태로 판단된다. 또한 Whittaker (1965)는 우점도 0.9 이상일 때는 1종이, 0.3~0.7일 때는 2~3종이, 0.3 이하 일 때는 다수의 종이 우점을 이룬다고 하였는데 개비자나무 집단의 경우 우점도가 0.067~0.346으로 다수의 종이 균일한 상태를 이루고 있어 임분의 성숙도가 높은 것으로 판단된다.
최대종다양성(H’ max)와 종다양성 지수(H’) 간의 값이 근접할수록 군집의 구조는 안정적이고, 값이 멀어질수록 구조가 불안정한데(Pielou, 1975), 개비자나무 군집구조는 다른 집단에 비해 적보산, 지리산, 두륜산에서 더 안정적인 것으로 나타났다.
산림군집의 구조적인 복잡성, 외부 교란 요인으로 부터의 안정성, 산림 발달 과정상의 성숙도는 종다양성과 정비례 하는 경향이 있다(Odum, 1969; Bazzaz, 1979). 개비자나무의 높은 종다양성 지수, 여러 종이 우점하는 군집형태는 천이진행 과정에서 외부교란의 요인으로부터 비교적 안정적이라 할 수 있으나 개비자나무의 종자 결실주기가 길고, 자웅이주이며, 서식지 단절화가 이루어지고 있어(Fu et al., 1999) 보전적 측면에서 지속적인 관리가 필요한 것으로 판단된다.
유사도지수
Table 6에서는 조사지간 종구성상의 유사한 정도를 나타내는 측도로서 유사도지수를 나타내고 있다.
유사도지수가 각 군집 간 20% 미만일 경우에는 이질적인 집단이고, 80% 이상일 때는 서로 동질적인 집단으로 간주한다고 하였다(Cox, 1972). 결국 생태적으로 유사한 집단의 유사도지수는 높게 나타나는 것으로, 6개 조사지를 대상으로 유사도지수(S,I)를 분석한 결과 지리산-두륜산이 39.4%, 지리산-백운산이 37.3%, 적보산-보개산이 35.7% 순으로 비교적 높게 나타났으나 80% 이상으로 동질성을 갖는 집단은 발견되지 않았다. 금원산-백운산이 12.7%, 적보산-금원산이 18.7%, 적보산-백운산 17.0%, 적보산-두륜산 19.8%, 금원산-지리산 19.1%으로 유사도지수 20% 이하를 나타내 이질적인 집단으로 나타났다.
Table 6. Similarity indices in the Cephalotaxus koreana habitats
| Habitatsz | C1 | C2 | C3 | C4 | C5 |
| C2 | 35.7 | ||||
| C3 | 18.7 | 28.5 | |||
| C4 | 22.5 | 30.4 | 19.1 | ||
| C5 | 17.0 | 29.6 | 12.7 | 37.3 | |
| C6 | 19.8 | 33.8 | 21.1 | 39.4 | 20.4 |
지리산-두륜산, 지리산-백운산 조사지 간 유사도지수가 비교적 높게 나타난 것은 남쪽에 위치하여 비슷한 서식환경을 가지며 그로인해 출현하는 종과 평균중요치값이 유사하기 때문인 것으로 판단되며, 금원산-백운산은 출현 종의 차이로 인해 유사도가 떨어진 것으로 판단된다.
유집분석에 의한 군집구조 분석
조사지역간 종구성에 대한 유사성을 분석하기 위해 6개 조사집단에서 출연한 수종 중 평균상대중요치 5% 이상을 나타내는 수종들을 Ward의 방법을 이용하여 Cluster분석을 실시하였다(Fig. 2).
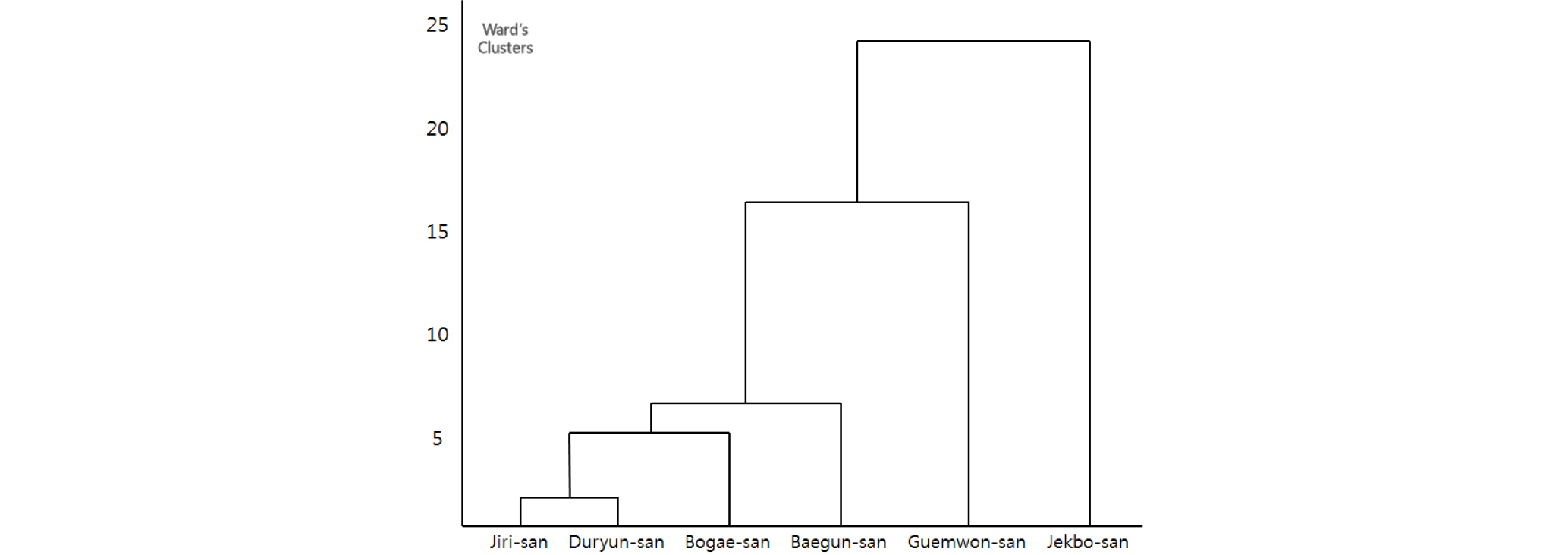
개비자나무 집단은 5개의 군집으로 분류되었으며, 지리산과 두륜산이 하나의 그룹을 이뤘을 뿐, 나머지 4개의 집단은 계단상의 형태를 보이고, 인접 집단 간의 구분이 명확하지 않아, 조사 집단 간 군집구조의 차이가 있음이 분석되었다. 결국, 개비자나무 자생지는 지역별로 우점하는 종이 서로 다르고, 다양한 환경을 갖는 것으로 판단되며, 이는 낮은 유사도지수를 나타낸 결과와 유사한 경향을 보였다.
상관관계분석
조사지별 개비자나무 생육형태를 분석하기 위해 환경요인과 토양분석값 및 식생조사 결과 간의 상관관계분석을 실시하였다(Table 7). 환경요인 중 상대습도와 교목층의 흉고단면적과 정의 상관관계를 형성하여 상대습도가 높을수록 생육이 좋은 것으로 나타났다. 또한 pH와 양이온치환용량(CEC) 함량 간에는 부의 상관관계(–0.842)를 형성하였고, 유기물함량과 밀접하게 관련있는 인자인 전질소, 양이온치환용량 값은 유기물함량과 정의 상관관계를 나타냈으나 개비자나무 생육에 직접적인 상관을 나타내는 인자는 발견되지 않았다.
Table 7. Correlation coefficients among environmental factors, structural properties and soil characteristic in Cephalotaxus koreana habitats
개비자나무 서식지의 종구성과 수고, 직경, 상대우점치의 상관관계를 분석한 결과 개비자나무의 수고는 교목층의 종다양도, 종다양도, 균재도와 정의 상관관계, 우점도와 부의 상관관계를 가지는 것으로 나타났다. 개비자나무의 직경은 종다양도와 정의 상관관계를 나타냈으며, 상대우점치 또한 교목층 종다양도와 정의 상관을 나타냈다. 이상의 결과를 종합하면 개비자나무는 종다양도와 균재도가 높고 임분의 성숙도가 높은 곳의 하층부에서 생육이 왕성한 것으로 나타났다. 이는 상층수목이 분포함에 따라 빛의 노출에 약한 개비자나무에게 유리한 생육지를 만듦으로써 생장을 발달시킨다고 보고한 선행연구 결과와 유사하였다(Fu et al., 1999).
적 요
본 연구는 우리나라에 자생하고 있는 개비자나무 6개 자연집단을 대상으로 군집구조를 분석하고 생육환경을 구명하여 합리적인 유지관리 방안의 기초자료를 제공하기 위해 실시되었다. 개비자나무 자생지는 온난대 낙엽활엽수림에 속하며 해발고 148~835 m, 경사 12~32°로 나타났다. 토양의 이화학적 특성을 분석한 결과 토양 산도는 pH 4.7~5.9로 약산성을 나타냈고, 유기물함량 5.72~15.99%, 양이온치환용량 14.1~19.9 cmolc/㎏-1로 유기물 유입이 활발한 것으로 확인되었으며, 치환성양이온 K⁺, Ca2+, Mg2+ 함량은 각각 0.25~0.48 cmolc/㎏, 0.79~6.68 cmolc/㎏, 0.31~1.73 cmolc/㎏으로 나타났다. 식생조사 결과 상층식생 우점종은 충주 적보산에서 떡갈나무, 쪽동백나무, 괴산 보개산에서 고로쇠나무, 느티나무, 거창 금원산에서 물푸레나무, 당단풍나무, 구례 지리산에서 졸참나무, 잔털벚나무, 광양 백운산에서 느티나무, 갈참나무, 해남 두륜산에서 상수리나무, 신갈나무로 집단 간 공통으로 우점하는 수종은 발견되지 않았고, 낙엽활엽수 중 참나무류가 우점하여 피음도가 높은 계곡부에 서식하는 것으로 조사되었다. 종다양도는 0.854~1.234, 균재도는 0.654~0.933, 우점도는 0.067~0.346으로 나타나 다수의 종이 균일한 상태를 이루는 성숙한 임분으로 나타났다. 종구성 및 환경요인과 개비자나무의 생육에 대한 상관관계를 조사한 결과 종다양도와 균재도는 개비자나무의 수고, 직경 및 우점도와 정의 상관관계를 가지는 것으로 나타나 성숙한 임분과 상층수목의 분포가 개비자나무에게 유리한 생육지를 제공하는 것으로 나타냈다.
