

서 언
재료 및 방법
억새 교배재료 출수기 단축을 위한 단일처리
억새 종별 일중 화분 발아시간 조사
억새 교배시 격리, 절단 화분친 이삭의 개화, 화분비산, 발아 지속방법 구명
억새 종별 자가수정 여부 구명
억새 종간교배 방법 효과 확인을 위한 시험교배
결과 및 고찰
단일처리에 의한 억새 출수 소요일수 조사
억새 종별 화분 발아시간
억새 교배시 절단 화분친 이삭의 개화, 화분비산 및 발아 지속
억새 종별 자가수정 여부
억새 종간교배 방법 효과 확인을 위한 시험교배
적 요
서 언
억새(Miscanthus species)는 C4 광합성 경로를 갖는 영년생 식물로 바이오매스 수량이 많고 양분이용 효율이 높아 유망한 바이오에너지작물이다(Arnoult and Brancourt-Hulmel, 2015; Greef et al., 1997). 동아시아와 태평양 제도가 원산으로서 약 17종의 억새가 알려져 있는데, 이 중 바이오매스 수량이 많은 것은 4배체 물억새(Miscanthus sacchariflorus), 2배체 참억새(Miscanthus sinensis) 그리고 이둘 두 종의 교잡종인 이질 3배체 억새(Miscanthus x giganteous)이다(Deuter et al., 1998; Moon et al., 2013; Greef et al., 1997). 한국은 억새 원산지 국가 중 하나로 강 또는 호수주변 제방에는 물억새, 언덕 등 마른 땅에는 참억새가 자생한다(Lee, 2002). 한국에서 2009년 물억새 유전자원 중 줄기가 크고 굵어 바이오매스 수량이 많은 ‘거대1호’를 선발하였다(Moon et al., 2010). 그러나 거대1호와 같은 종자가 맺히는 품종은 종자가 바람에 의해 널리 확산되므로 유럽, 북미 등 억새 원산지가 아닌 지역에서는 생태계 교란 방지를 위해 재배를 규제되고 있다(Barney and DiTomaso, 2008; Bonin et al., 2014; Quinn et al., 2010). 이러한 국가들은 바이오매스 생산을 위해 불임성인 이질 3배체 억새(M. x giganteus)을 재배한다. 이 종은 1930년대에 일본 남부지역에서 수집되어 유럽으로 전래된 것으로 동일 영양체에서 유래된 단일 품종이다(Greef et. al., 1997; Lewandowski et al., 2000). 따라서 기후환경이 다른 지역에서는 품종 고유 특성을 발휘하기 어렵고, 넓은 면적에 집단적으로 재배할 경우 돌발 병해충으로 재배단지 전체가 사멸할 위험이 있다(Beccari et al., 2010; Nishiwaki et al., 2011). 이러한 위험성을 회피하기 위해 다양한 품종의 이질 3배체 억새를 개발할 필요가 있으나 연구역사가 짧고 적절한 육종방법이 정립되지 않아 우량품종 개발 성과가 미흡한 실정이다(Deuter et al., 1998). Deuter (2009)는 폐쇄된 유리온실 내에서 종자친으로 4배체 물억새 수집종 1종과 화분친으로 2배체 참억새 여러 수집종의 집단 재배로 무작위 다교배(poly cross)를 유도하여 몇 계통의 종간 교잡종을 얻었다. 이러한 다교배로 육성한 종간교잡종의 종자친은 수집종 1종으로 그 특성을 알 수 있으나 화분친을 알기 어려워 형질이 우수한 종자친과 화분친으로 교배조합을 작성하기 어렵다. 벼 등 육종연구 역사가 오래된 작물은 대부분의 우수 교배재료의 출수․개화기가 조사되었고, 제웅, 화분 비산시간 등을 고려한 교배 및 격리 등 교배 방법이 정립되어 우량품종 육성을 위해 단교배(single cross)를 이용하고 있다. 국내에서 이질 3배체 억새 품종 육성을 위해 물억새와 참억새와 개화 패턴을 조사하여 물억새가 참억새에 비해 개화기가 늦는 것을 밝혔으나(An et al., 2015), 우수 화분친과 종자친 양성 방법, 개화기를 조절하는 방법, 교배 및 격리방법 등 단교배 필수요소에 관한 연구결과가 없는 실정이다. 본 연구는 비 원산지 국가에서 재배 가능한 이질 3배체 억새 육종법 개발을 위해 물억새와 참억새 단교배에 필요한 교배재료의 일장감응, 화분 비산 및 발아특성, 교배 및 격리 방법 등 단교배 조건을 구명하기 위해 수행하였다.
재료 및 방법
억새 교배재료 출수기 단축을 위한 단일처리
시험 재료는 전남 무안군 청계면 소재 바이오에너지작물연구소 억새 유전자원 보존포장(34.97 °N, 126.45 °E)에서 굴취하였다. 4배체 물억새는 수량은 많지만 출수기가 늦은 BM 00240 (거대1호) 등 수집종 3종을, 2배체 참억새는 BM 00830 등 수집종 3종을 사용하였다. 억새의 양분 저장 및 번식기관은 지하 5~15 ㎝에 발달하는 지하경(rhizome)으로, 물억새는 연간 20~100 ㎝ 정도 직선형으로 길게 신장하지만 참억새 지하경은 만곡형으로 연간 신장 길이가 10~15 ㎝ 정도로 짧다(Barney and DiTomaso, 2008; Lee, 2002). 이러한 종별 지하경 신장 특성 때문에 4배체 물억새 지하경은 전년도 신장한 것을 4월 상순에 굴취 후 길이가 40 ㎝ 이상인 것을 사용하였다. 2배체 참억새는 시험 1년 전 10 ㎝ 길이 지하경을 굴취하여 4 L 와그너포트에 1년간 양성한 것을 사용하였다. 시험재료 재배를 위한 상토는 코코피트, 피트모스, 제오라이트, 질석, 도로마이트가 각각 47.2, 35.0, 7.0, 10.0, 0.6%가 함유된 원예용 상토(FarmHanong Ltd, Korea)를 준비하였다. 직경과 높이가 모두 40 ㎝인 대형 포트에 상토를 채우고 포트 1개당 지하경 4개씩 재식하였다. 억새를 재식한 포트는 6월 31일까지 자연일장 조건에서 재배한 후, 7월 1일부터 수집종별 5개 포트씩 12시간 일장 조건에서 재배하였고, 다른 5개 포트는 계속 자연일장 하에서 재배하였다. 출수 소요 일수는 지하경에서 새싹이 출현한 날짜를 맹아기, 이삭이 지엽을 뚫고 나온 날짜를 출수기로 하여 맹아기에서 출수기까지 소요되는 일수로 하였다. 조사결과는 SAS 통계분석 패키지(version 9.3-2012; SAS Institute, Cary, NC, USA)로 스튜던트 t-분석을 수행하여 유의성을 검정하였다.
억새 종별 일중 화분 발아시간 조사
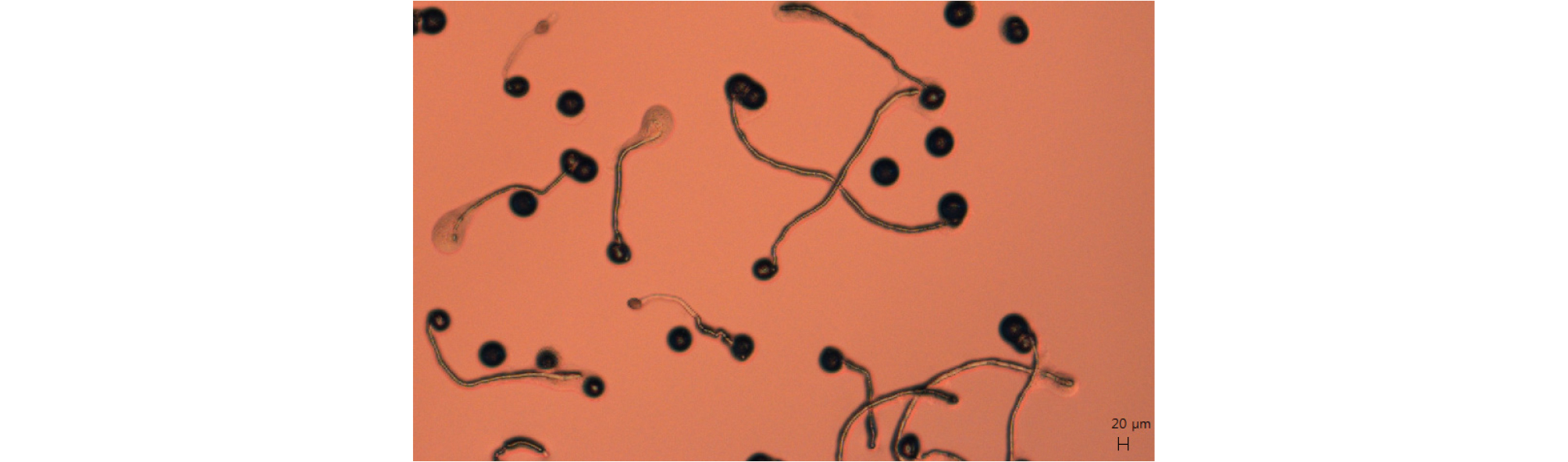
화분발아 배지로 10 g.L-1 sucrose가 첨가된 한천배지를 직경 60 ㎜ 패트리디쉬에 채워 사용하였다. 10월 상순 오전 6시부터 9시 까지 1시간 간격으로 유전자원 보존 포장에서 출수된 물억새와 참억새 이삭의 화분을 준비된 패트리디쉬에 털어 접종하였다. 화분이 접종된 패트리디쉬는 25℃ 배양기에 3시간 배양 후 100배 현미경(Carl zeiss DE/Axio Scope A1, Germany)으로 관찰하였다. 화분 발아율은 현미경 관찰 시 패트리디쉬를 종, 횡으로 옮기면서 현미경에 장착된 카메라(Axio Cam IC 1, Germany)로 100배로 촬영하여 50∼100개 화분 중 Fig. 1과 같이 발아관이 신장된 화분의 백분율을 조사하였다.
억새 교배시 격리, 절단 화분친 이삭의 개화, 화분비산, 발아 지속방법 구명
억새 교배 격리공간은 직경과 길이가 각각 20 ㎜, 170 ㎝인 철제 파이프 4개와 5 ㎜ 두께 강철제 고리(Ring) 2개로 직경과 높이가 각각 73 ㎝, 160 ㎝인 고정틀을 설치하고 동일 크기의 백색 부직포 원통을 제작하여 고정틀에 씌워 준비하였다. 백색 부직포는 최대 구멍 크기가 약 12 ㎛로 조기 출수한 참억새 유전자원(BM 00938)의 화분 직경(13.6~17.4 ㎛)에 비해 작은 것을 확인하였다(Fig. 2). 이때 화분 및 부직포 구멍 크기 측정을 위해 100배 현미경(Carl zeiss DE/Axio Scope A1, Germany), 카메라(Axio Cam IC 1, Germany) 및 크기측정 프로그램(IMAGE-PRO Express, Germany)을 사용하였다.

화분친 이삭 보존액은 증류수 1 L에 sucrose 100 g, 질산은(AgNO3) 250 ㎎을 녹인 후 1 N H2SO4로 pH를 3.5로 조절하였다. 화분친 이삭 보존 용기는 polyethylene 원심분리 튜브를 활용하여 직경과 길이가 각각 3 ㎝, 30 ㎝ 크기로 제작 후 햇볕투과와 온도상승 방지를 위해 알루미늄 호일로 감쌌다. 준비된 용기에 보존액을 50, 100, 150 mL 씩 채우고, 대조구로 보존액을 채우지 않은 빈 용기를 준비하였다. 물억새와 참억새 유전자원 BM 00184, BM 00558 이삭을 출수 직후 화분이 비산되기 전에 지엽 포함 상위엽 3매가 달리도록 약 140 ㎝ 길이로 절단하였다. 절단 이삭은 화분친 이삭 보존액이 채워진 용기에 2개씩 꽂아 설치된 백색 부직포 격리 공간 내에 고정시켰다. 보존액량별 화분친 이삭 개화 및 화분비산 지속 여부는 매일 오전 7시경에 달관조사하였다. 시험 기간 중 화분 발아율은 화분비산이 오래 지속될 것으로 짐작되는 150 mL 보존액에 꽂은 이삭을 대상으로 상기 ‘억새 종별 화분 비산 및 발아시간 조사’ 방법으로 조사하였다. 시험 기간 중 격리공간과 노지의 최고온도는 temperature logger (TGP-4017 Tinytag, England)로 측정하였다.
억새 종별 자가수정 여부 구명
시험재료로 유전자원 보존포장에 재식된 참억새(M. sinensis) BM 00766 등 수집종 5종, 물억새(M. sacchariflorus) BM 00240 등 수집종 5종을 사용하였다. 상기 억새는 유전자원 보존용 매립 PVC 통(L: 102 ㎝ × W: 72 ㎝ × H: 35 ㎝, 바닥을 오려낸 것)에 식재된 것으로 줄기를 50%씩 분할하여 자가수정, 자연교배에 사용하였다. 자가수정은 출수 직후 상기 ‘교배 격리공간’ 설치와 같은 방법으로 고정틀을 설치 후 백색 부직포 원통을 씌웠고, 나머지 50%는 유전자원 보존포장의 다른 수집종들과 자연교배가 되도록 외부에 노출시켰다(Fig. 3). 유전자원 보존포장에는 전국 각지에서 수집된 물억새와 참억새 유전자원 1,200여 수집종이 식재되어 자연교배가 용이한 조건이었고(An et al., 2015), 전년도에 자연방임 상태에서 임실을 확인하였다. 처리 30일 후 격리용 부직포 원통을 제거하고, 40일 후 이삭을 수확 후 3개 소지경을 추출하여 전체 영(穎) 중의 임실된 것을 계수, 백분율로 자가수정율과 자연교잡 임실율을 조사하였다. 억새 교배 및 임실 기간 중 평균기온, 최고기온, 최저기온은 시험장소인 바이오에너지작물연구소에 설치된 농촌진흥청 농업기상서비스 농업기상관측시스템(RDA, Korea)의 농업기상관측 자료를 활용하였다.

억새 종간교배 방법 효과 확인을 위한 시험교배
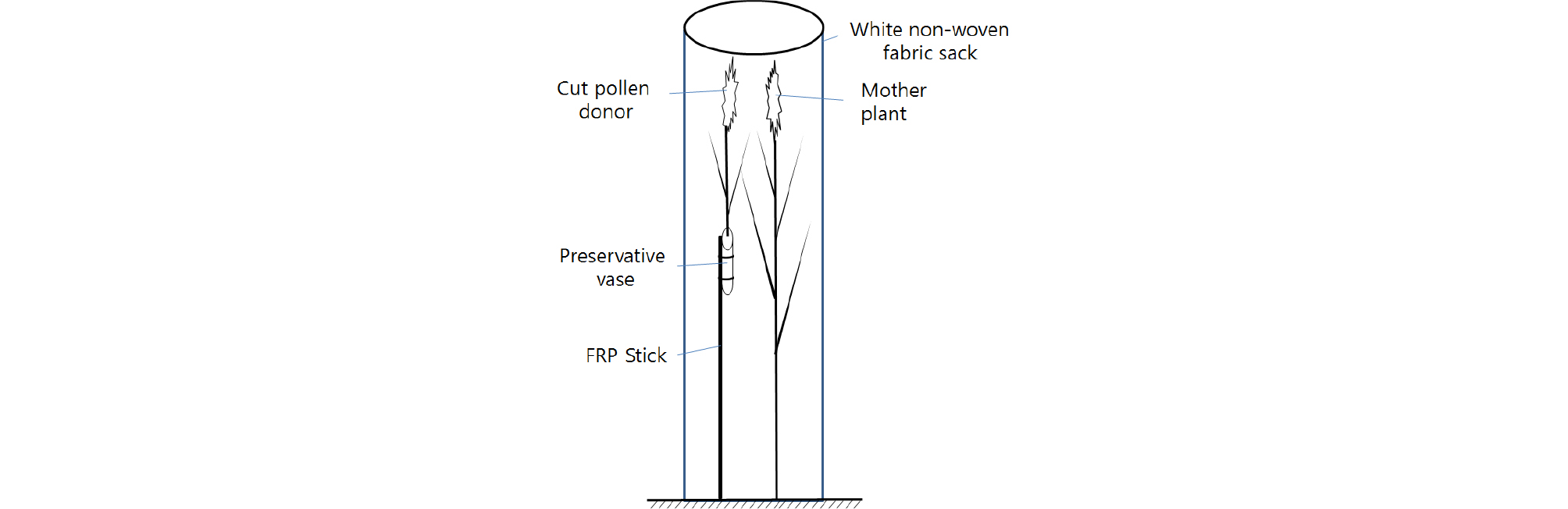
종자친은 물억새 4배체 유전자원 BM 00240과 BM 00242를, 화분친은 2배체 참억새 BM 00677등 수집종 7종으로 총 14조합을 교배하였다. 물억새 4배체는 출수기 단축 시험에서 단일처리로 자연일장 재배 억새에 비해 일찍 출수한 것이었고, 참억새 2배체는 유전자원 포장에 재식된 것 중에서 종자친과 동일한 시기에 출수한 것이었다. 격리공간은 10월 상순경 종자친 포트 주위에 4 m 철제 파이프(∮20 ㎜) 4개를 50 × 50 ㎝ 간격으로 세우고 꼭대기와 중간에 강철제 활죽으로 만든 고리(∮70 ㎝)로 고정시킴으로써 확보하였다. 상기 ‘화분친 이삭 보존 방법’과 동일하게 개화 직전의 이삭을 절단하여 절화 보존액에 꽂은 후 ∮8 ㎜ FRP 막대로 종자친 이삭 높이에 맞도록 절화 보존액 용기에 고정시켰다(Fig. 4). 격리는 동일 실험에서 확인된 백색 부직포로 ∮73 ㎝, 높이 4 m 원통을 제작하여 격리공간 지지틀에 1개월 동안 씌웠다. 11월 하순경 이삭을 수확하여 교배조합별로 이삭 3개의 임실종자 수를 조사하여 SAS 프로그램 9.2 버전(Statistical Analysis System ver. 9.2, SAS Institute, Cary, NC, USA)으로 Duncan 다중검정을 수행하였다.
결과 및 고찰
단일처리에 의한 억새 출수 소요일수 조사
자연일장과 단일 조건에서 재배한 억새의 출수 수요일수 차이를 조사한 결과는 Table 1과 같다. 참억새와 물억새 모두 7월 1일 이후 12시간 일장의 단일조건에서 재배한 것이 자연일장에 재배한 것에 비해 출수 소요일수가 17~29일 정도 단축되어 자연일장에 재배한 것과 유의적인 차이를 보였다(p ≤ 0.001). 억새는 단일성 식물로 여름철 일장이 긴 고위도 지역에 적응한 유전자원은 빨리 출수되고, 저위도 지역에 적응한 것은 늦게 출수하는 경향이 있다(An et al., 2015). 이렇게 늦게 출수한 유전자원은 영양 생장 기간이 길어 바이오매스 수량이 많으므로 육종재료로 가치가 높다(Deuter et al., 1998). 그러나 늦게 출수한 유전자원은 교배 후 종자 등숙기간 중 온도가 낮아 종자가 맺히지 않을 수 있다(Table 1).
Table 1. Difference of days to heading between natural and short day length in M. sacchariflorus and diploid M. sinensis
y*** Indicate statistical significance at p < 0.001 from Student's t-tests comparing days to heading between natural and short day length.
본 시험 결과 억새 육종재료를 자연일장에서 재배하면 출수기가 늦어 교배 후 짧은 등숙기간으로 인해 종자가 맺히지 않을 수 있지만 단일조건 재배로 출수기를 단축하면 교배 후 등숙기간이 충실한 종자를 얻을 수 있을 것으로 판단된다.
억새 종별 화분 발아시간
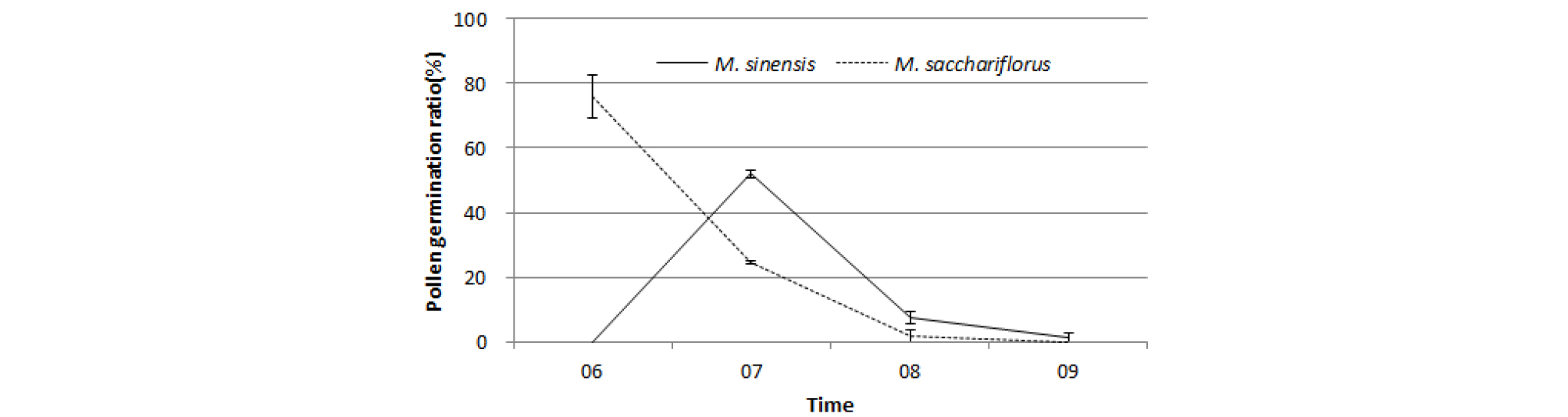
물억새와 참억새의 하루 중 시간별 화분발아율을 조사한 결과는 Fig. 5와 같다. 물억새는 오전 6시에 화분 발아가 왕성하였으나 시간이 경과할수록 낮아져 오전 8시에는 발아율이 10% 이하였다. 참억새 화분은 오전 6시에는 발아하지 않았으나 오전 7시에 50% 이상이 발아하였고 8시에는 물억새와 같이 발아율이 10% 이하였다. 바이오에너지용 3배체 종간교잡종인 M. x giganteous는 종자친이 4배체 물억새이고, 화분친이 2배체 참억새로 알려져 있다(Arnoult and Brancourt-Hulmel, 2015; Clifton-Brown et al., 2008; Deuter et al., 1998; Moon et al., 2010). 억새는 바람으로 화분이 매개되는 풍매화(Moon et al., 2013; Nishiwaki et al, 2011; Uwatoko et al., 2016)이기 때문에 3배체 종간교잡 품종을 육성하려면 참억새 화분 활력이 가장 좋은 오전 7시경 화분친 이삭을 흔들기 등으로 개방된 수술에서 화분 비산을 조장할 필요가 있다고 사료된다.
억새 교배시 절단 화분친 이삭의 개화, 화분비산 및 발아 지속
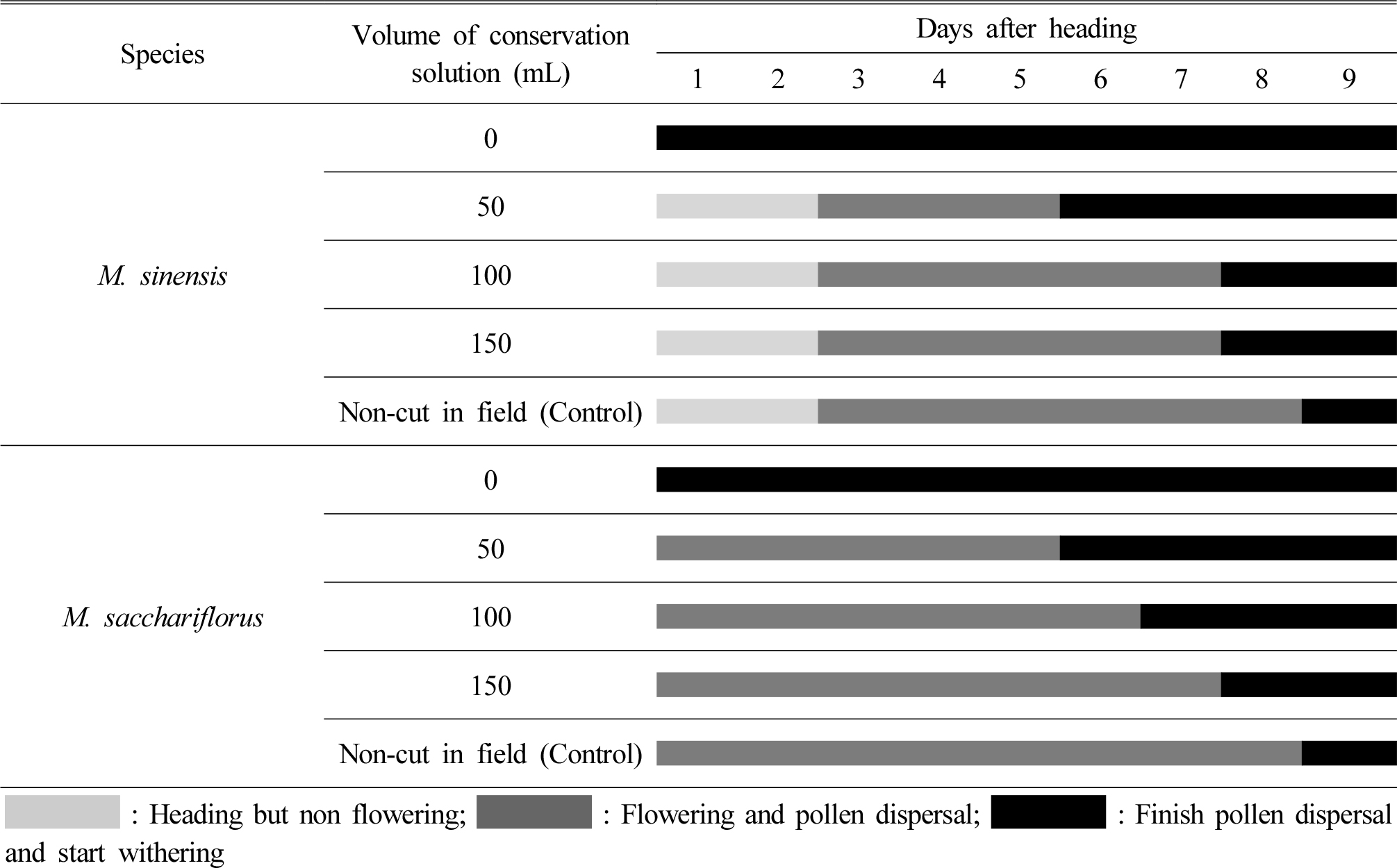
절화 보존액 용량별 이삭의 개화 및 화분비산 지속기간을 관찰한 결과는 Fig. 6과 같다. 절화 보존액량이 많을수록 절단한 이삭의 활력 유지 일수가 증가하여 150 mL에 꽂은 물억새 이삭은 7일간 개화 및 화분비산을 지속하였고, 참억새 이삭은 2일 경과 후 화분비산을 시작하여 7일 까지 지속하여 절단하지 않은 이삭의 개화 및 화분 비산 기간 8일과 큰 차이가 없었다. 절화 보존액이 없는 빈 용기에 꽂은 이삭은 억새 종에 관계없이 1일 후 고사하여 화분을 비산하지 않았다. 식물의 인공교배시 원치 않는 화분 혼입을 방지하면서 화분친의 활력을 지속시키는 것이 중요하다(Kim et al., 2015). 절단된 억새 이삭과 비슷한 관상용 절화는 활력을 지속시키기 위해 절화 보존액으로 이삭에 양분과 수분을 지속적으로 공급해 주어야 한다(Hwang et al., 2009). 본 실험 결과로 절단한 억새 이삭을 150 mL의 절화 보존액에 꽂으면 최대 7일 까지 활력이 유지되어 화분을 비산시킬 수 있다. 따라서 이질 3배체 육성을 위한 종간교배시 갓 출수한 물억새 이삭(종자친) 곁에 출수 2일이 경과하여 개화 직전인 참억새 이삭(화분친)을 절단하여 150 mL의 절화보존액에 꽂은 다음 모본 이삭 높이와 동일하게 고정 후 격리하는 것이 좋을 것으로 판단된다.
억새 이삭을 절화 보존액에 꽂아 부직포로 격리한 공간에 정치한 후 이삭에서 비산된 화분의 날짜 경과에 따른 발아율 변화를 조사한 결과는 Fig. 7과 같다. 참억새 화분 발아율은 처리 4일 후 까지 40% 이상을 유지하였고 5일 후에는 26%로 낮아져 절화 보존액에 꽂아 백색 부직포로 격리한 이삭과 노지 이삭의 화분의 발아율과 비슷하였다. 물억새도 참억새와 유사하게 절화 보존액에 꽂아 백색 부직포로 격리한 이삭과 노지 이삭 화분 발아율이 비슷하면서 처리 4일 후에 40%를 유지하였다. 실험 기간 중 백색 부직포 격리공간내 최고온도는 노지에 비해 높았으나 35℃ 이하를 유지하였다(Fig. 8).
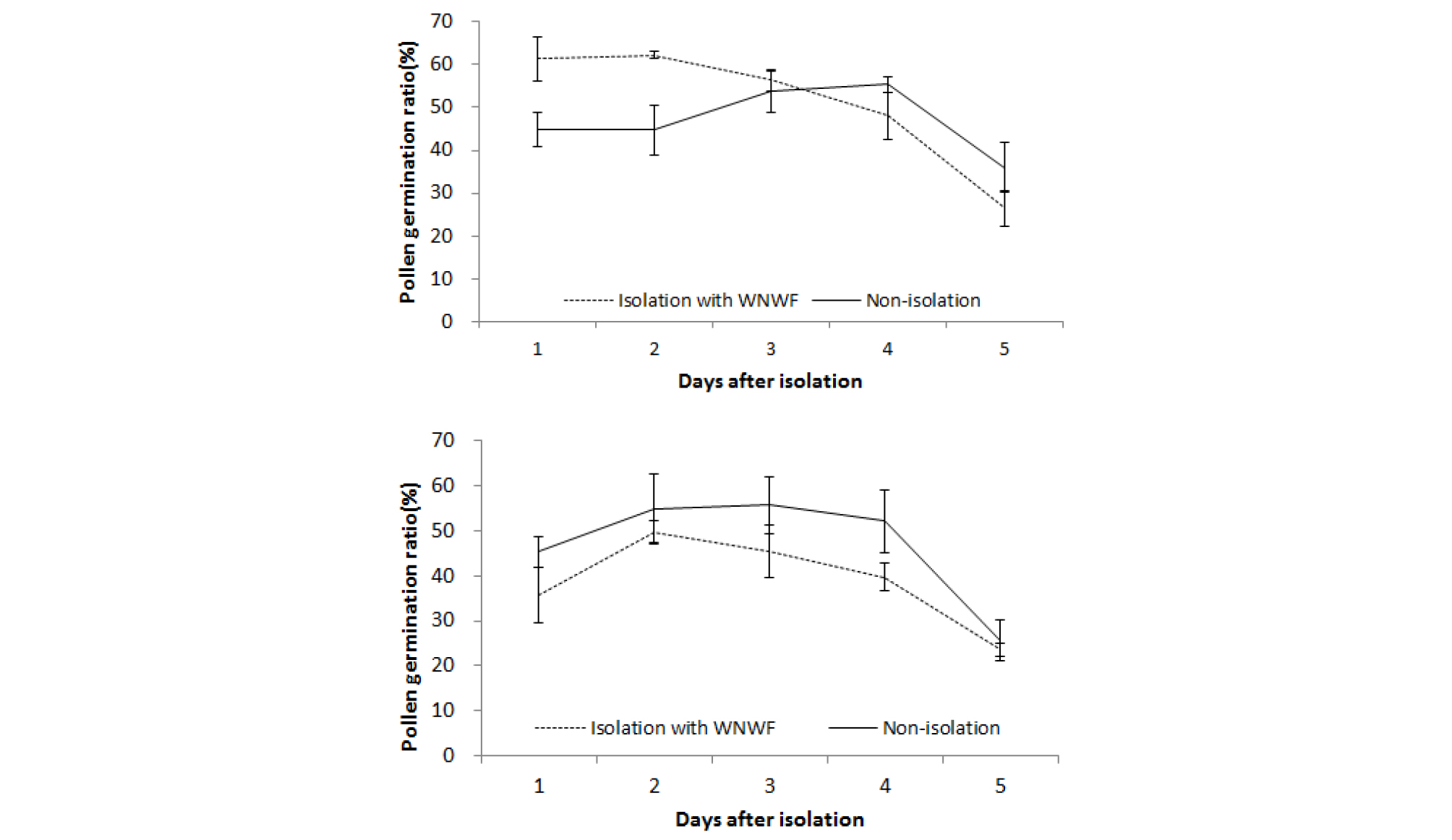
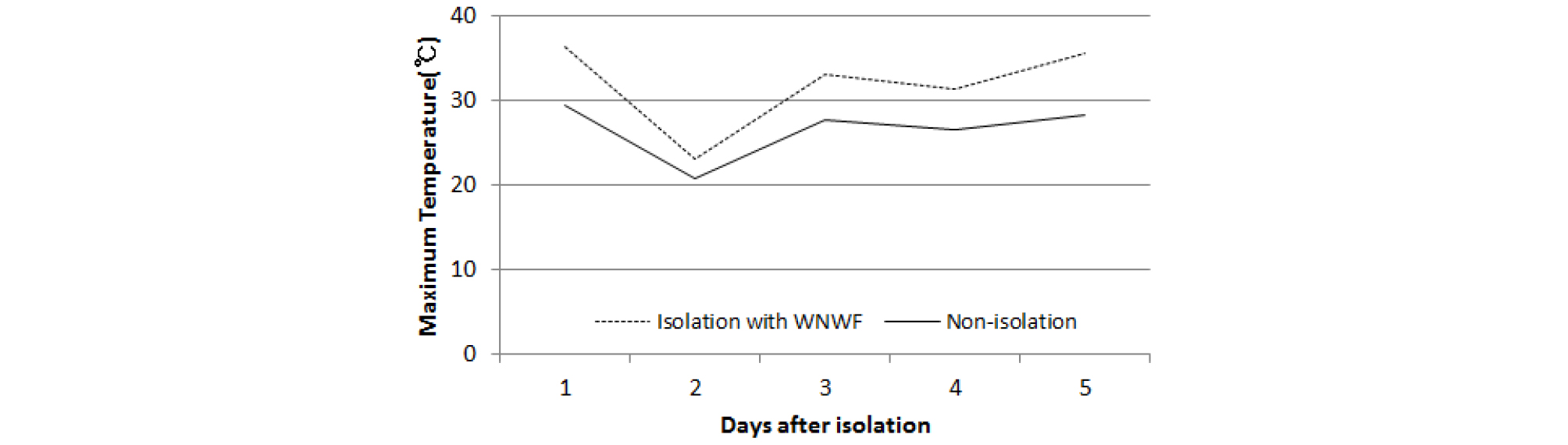
수분용 복숭아 화분은 20~25℃에서 화분 발아율이 80% 이상을 유지하므로 수분을 위한 최적 온도이지만, 온도가 35℃로 상승하더라도 56%의 발아율이 유지되었다(Cheon et al., 2006). 인공수분으로 착과율을 높여야만 하는 복숭아와는 달리 억새 종간교잡은 영년생 잡종 식물체을 얻는 것이 목적이므로 40%의 화분발아율도 충분할 것으로 판단된다. 본 실험 결과 격리공간 온도를 35℃ 이하로 유지한다면 화분 활력이 크게 감소하지 않을 것으로 보인다. 불임성 이질 3배체 억새는 화분친이 참억새로 알려져 있으므로 종간교배시 부직포로 격리한다면 절화보존액에 꽂은 화분친 이삭의 화분 발아력은 4일 후까지 유지될 것으로 기대된다.
억새 종별 자가수정 여부
억새 수집종의 백색 부직포 격리를 통한 종별 자가수정율과 자연교잡율을 조사한 결과는 Table 2와 같다. 참억새와 물억새의 자가수정율 범위는 각각 1.0~2.5%, 0.0~1.2% 수준으로 낮았다. 본 실험 결과, 억새는 자가불화합성 식물이므로 품종육종을 위한 인공교배시 종자친 이삭의 수술을 제거할 필요가 없다는 여러 선행연구 결과와 일치하였다(Clifton-Brown et al., 2008; Deuter et al., 1998; Greef et al., 1997).
Table 2. Ratio of seed set on spike of Miscanthus species by isolationz for self-fertilization, and non-isolation for natural fertilization
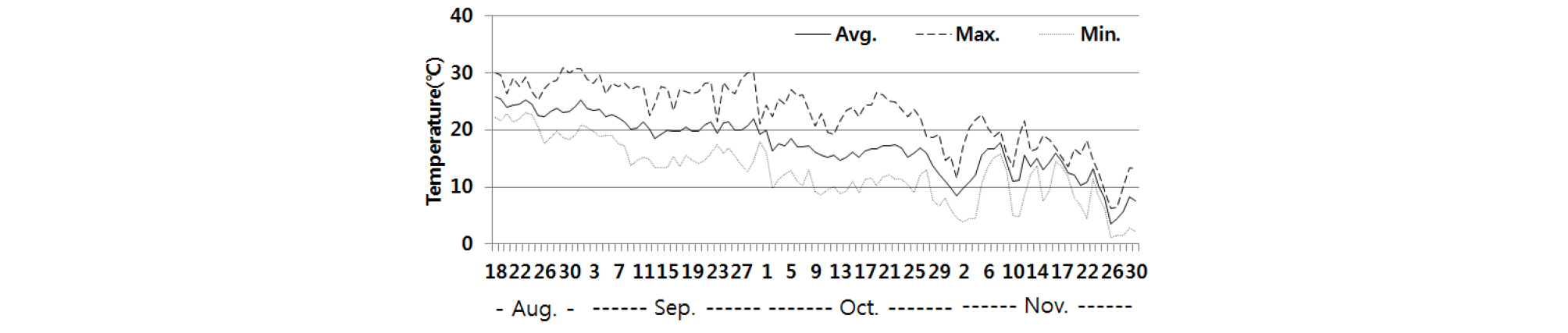
자연교잡율의 경우 참억새는 20.8~54.4%로 비교적 높았고, 물억새는 0.0~53.9%로 수집종별 차이가 컸다. 일반적으로 식물은 수정되면 일정 수준의 온도를 유지해야만 종자가 등숙이 되는데, 억새와 같이 단자엽 식물인 벼는 출수 후 40일간 21~23℃의 평균기온이 유지되어야 종자가 등숙한다(Yun and Lee, 2001). 참억새의 자연교잡 임실율이 높은 것은 출수기가 8월 중하순으로 교잡 후 9월 하순~10월 상순까지 평균기온이 15℃ 이상을 유지하였기 때문인 것으로 보인다(Fig. 9). 물억새에서는 9월 5일 이전에 출수한 BM 01011과 BM 01013은 임실율이 각각 53.9%, 33.1%였으나 출수일이 9월 10일인 BM 01009는 6.3%로 낮았고, 출수기가 9월 27일인 BM 00240과 BM 00242는 등숙된 종자가 없었다. 이것은 Fig. 9에서 보는바와 같이 10월 하순과 11월 상순에 최저기온이 10℃ 이하로 급격히 낮아져 종자가 등숙되지 않았기 때문인 것으로 보인다. 억새는 출수기가 늦을수록 영양생장량이 증가하여 바이오매스 수량이 많아진다(An et al., 2015; Lewandowski et al., 2000; Moon et al., 2010). 따라서 바이오에너지용 종간교잡 이질 3배체 억새 품종육성을 위해 수량이 많은 4배체 물억새를 종자친으로 활용하려면 출수기를 단축시킴으로서 수정 후 저온으로 낮아지기 전에 등숙시켜야 할 것으로 보인다.
억새 종간교배 방법 효과 확인을 위한 시험교배
억새 종간교배 방법 효과 확인을 위해 4배체 물억새 유전자원 BM 00240과 BM 00242를 모본으로, 2배체 참억새 유전자원 BM 00677 등 7종을 부본으로 14조합을 시험교배한 결과는 Table 3과 같다. 모본으로 BM 00240을 사용한 조합의 종자 수는 7∼28 seeds/ear로 BM 00242가 모본인 것의 2∼3 seeds/ear에 비해 많았다. 일반적으로 식물의 종간교배는 임실율이 낮고, 획득된 종자도 활력이 낮아 잡종 식물체를 육성하기 어렵다(Esen and Soost, 1971; Johnston et al., 1980; Kim et al., 2018). 이 연구 결과 교배 조건이 동일하더라도 모본에 따라 임실율이 차이가 있다고 판단된다. 이 시험 결과 얻은 교배종자 437립으로 유식물 323개체를 발아시켰고, 이 중 290개체가 잡종괴사(hybrid necrosis) 현상으로 고사되었지만 이질 3배체 억새(M. x giganteus) 12개체를 얻었다(Moon et al., 2018).
Table 3. Number of seed set by experimental crossing to evaluate new crossing methods for breeding of interspecific hybrids between tetraploid M. sacchariflorus and diploid M. sinensis
적 요
본 연구는 바이오에너지용 이질 3배체 억새(Miscanthus x giganteus) 품종육성 위한 교배재료인 물억새(M. sacchariflorus)와 참억새(M. sinensis) 출수기 단축과 단교배 조건을 구명하기 위해 수행하였다. 물억새와 참억새를 12시간 일장, 자연일장 조건에서 재배하여 출수기 단축에 미치는 단일효과를 조사하였다. 출수기에 일중 화분 발아시간, 절단한 화분친 이삭 활력 유지 및 격리방법 등 단교배 조건을 구명하여 이를 활용한 억새의 자가수정 여부를 조사하고 시험교배를 실시하였다. 참억새와 물억새 모두 12시간 일장의 단일조건에서 재배한 것이 자연일장에 재배한 것에 비해 출수 소요일수가 단축되었다. 화분 발아 시간은 물억새는 오전 6시에 왕성하였으나 시간이 경과할수록 발아율이 낮아져 오전 8시에는 10% 이하만 발아하였다. 참억새 화분은 오전 7시에 50% 이상이 발아하였고 그 이후로 낮아졌다. 화분친인 참억새 이삭을 절단하여, 절화 보존액에 꽂아 백색 부직포로 격리하였을 때 절화 보존액량이 많을수록 활력 유지 일수가 증가하여 150 mL에서 물억새, 참억새 모두 7일간 개화 및 화분 비산을 지속하였다. 이 때 화분 발아율은 참억새와 물억새 모두 4일까지 40%를 유지하였다. 참억새와 물억새는 자가수정율이 2.5% 이하로 낮고, 자연교잡 임실율은 출수기가 빠른 유전자원에서 54.4%까지 높았다. 억새 종간교배 방법 효과 확인을 위해 물억새 4배체와 참억새 2배체 간 14조합 시험교배 결과 총 437립의 교배종자를 얻었다. 본 연구로 도출된 억새 종간 단교배 방법은 우수한 종자친과 화분친으로 교배종자를 얻을 수 있어 향후 바이오매스 수량이 많으면서, 종자가 맺히지 않은 이질 3배체 품종육성에 활용할 수 있을 것으로 판단된다.
