

서언
재료 및 방법
실험재료
추출물 제조
세포 생육 억제활성 측정
SDS-PAGE 및 Western blot 분석
Reverse transcriptase-polymerase chain reaction (RT-PCR)
Transient transfection
통계분석
결과 및 고찰
TY-NS-B의 A549세포에 대한 세포생육억제와 β-catenin 단백질 수준 감소 유도 활성
TY-NS-B의 GSK3β 비 의존성 β-catenin 단백질 분해 유도활성
적요
서언
폐암은 전 세계적으로 암과 관련 사망의 주요한 원인으로, 국가적으로 건강관리시스템에 상당한 사회·경제적 영향을 미친다고 보고되고 있다(Siegel et al., 2018). 2018년 보고된 자료에 따르면, 한국에서 2018년 총 204,909건의 새로운 암이 발생하며, 전체 암으로 인한 사망률 중 폐암으로 인한 사망률이 1위를 차지한다(Jung et al., 2018). 소세포폐암(Small cell lung cancer)과 비교할 때, 비소세포성 폐암(Non-small cell lung carcinoma)은 폐암의 약 75∼85%를 차지한다(Eisenberg et al., 1998). 비소세포성 폐암은 폐암 치료요법으로 알려져 있는 방사선 치료와 화학치료에 대한 내성을 나타내어 일반적으로 수술을 통한 절제를 하기에는 늦은 단계에서 발생되어 5년 생존율이 15% 이하로 알려져 있다(Erridge et al., 2007). 비록 비소세포성 폐암의 조기 발견과 치료법이 개선되고 있지만, 비소세포성 폐암의 빠른 재발과 진행으로 전반적인 예후는 여전히 열악한 상태이다(Guessous et al., 2007). 그러므로 폐암치료를 위한 효과적인 치료제와 치료를 위한 새로운 전략이 시급히 필요한 실정이다(Lin et al., 2019).
약용식물 추출물이나 다양한 성분이 폐암을 포함한 여러 종류의 암에 대해 항암활성을 나타내는 것으로 보고되고 있어 최근 약용식물이 악성 종양 치료의 대안으로 주목 받고 있다(Eisenberg et al., 1998; Kim et al., 2017; Risberg et al., 1998; Sadava et al., 2002; Yin et al., 2004).
겨우살이는 교목이나 관목의 가지에 기생하는 단항목에 속하는 현화식물로서 우리나라에는 단향과의 겨우살이[Viscum coloratum (Komarov) Nakai f. coloratum], 붉은 겨우살이 [Viscum coloratum (Komarov) Nakai f. rubroaurantiacum (Makino) Kitagawa], 동백나무겨우살이 [Korthalsella japonica (Thunb.) Engl] 등 3분류군과 꼬리겨우살이과의 꼬리겨우살이(Loranthus tanakae Franch. et Sav.)와 참나무겨우살이[Taxillus yadoriki (Sieb. ex Maxim.) Danser] 등 2분류군을 합하여 2과 4속 5분류군이 분포하고 있다(Kim et al., 2013). 겨우살이는 항암, 항염증, 항HIV 및 면역조절과 같은 다양한 기능성을 가지고 있다고 보고되고 있다(Mengoni et al., 2002; Ovesna et al., 2004; Safayhi and Sailer, 1997; Weissenstein et al., 2011). 참나무겨우살이가 기주하는 기주식물[삼나무(Cryptomeria japonica), 참식나무(Neolitsea sericea), 벚나무(Prunus serrulata), 녹나무(Cinnamomum camphora) 및 상수리나무(Quercus acutissima)] 및 부위(잎과 가지) 별 항염증활성과 항암활성 연구에서, 기주식물별로는 참식나무에 기주하는 참나무겨우살이가 그리고 부위별로는 가지추출물이 잎추출물에 비해 항염증 및 항암활성이 가장 높았다(Park et al., 2018a; Park et al., 2018b). 참식나무 기주 참나무겨우살이 가지 추출물은 LPS에 의해 유도되는 NF-κB와 MAPK의 활성화를 억제하고, ATF3의 발현을 촉진하여 염증매개인자의 발현을 억제하였다(Park et al., 2018a). 그리고 참식나무 기주 참나무겨우살이 가지 추출물은 cyclin D1의 분해를 통해 대장암세포의 생육을 억제하였다(Park et al., 2018b).
본 연구에서는 참식나무 기주 참나무겨우살이 가지 추출물의 비소세포성 폐암에 대한 항암활성을 확인하기 위해, 비소세포성 폐암 세포인 A549에 대한 세포생육억제활성을 측정하였고, 그 조절 기전으로 β-catenin의 분해를 조사하였다.
재료 및 방법
실험재료
비소세포성 폐암세포의 배양을 위해 사용된 배지인 Dulbecco’s modified Eagle’s medium (DMEM)/F-12 1 : 1 modified medium (DMEM/F-12)는 Lonza (Morristown, NJ, USA)에서 구매하였다. 특이적 효소 억제제인 LiCl 및 MG132와 대장암 세포의 생육억제효능을 평가하기 위한 3-(4,5-dimethylthizaol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)는 Sigma-Aldrich (St. Louis, MO, USA)사에서 구매하였다. 그리고 western blot 분석을 위한 항체인 β-catenin, phospho-β-catenin, Flag 및 actin은 Cell Signaling Technology (Danvers, MA, USA)에서 구매되었다.
추출물 제조
본 실험에 사용된 참식나무 기주 참나무겨우살이 가지 시료는 경북 영주에 위치한 국립산림과학원 산림약용자원연구소로부터 제공받았다. 음건·세절한 시료 20 g을 400 mL의 에탄올을 가하여 상온에서 3일 동안 교반하면서 추출하고, 원심분리하여 상등액을 회수하였다. 회수된 상등액은 40℃ 이하의 중탕에서 감압 환류 냉각장치(N-1110S, EYELA, Tokyo, Japan)로 감압농축 그리고 동결건조되었다. 동결건조된 추출물은 -80℃의 냉동고에 보관하면서 항암활성 분석을 위한 시료로 사용되었으며, 추출물(TY-NS-B)은 dimethyl sulfoxide (DMSO)에 용해하여 실험에 사용하였다. 대조군은 DMSO를 0.1%를 초과하지 않는 범위에서 사용하였다.
세포 생육 억제활성 측정
본 연구에서 사용된 비소세포성 폐암 세포인 A549는 American Type Culture Collection (Manassas, VA, USA)에서 구매되었고, 세포는 10% fatal bovine serum, 100 U/mL penicillin과 100 ㎍/mL streptomycin이 포함된 DMEM/F-12 배지로 37℃, 5% CO2 하에서 배양되었다. 비소세포성 폐암 세포인 A549의 생육억제는 MTT assay로 측정하였다. A549세포를 96-well plate에 well 당 1 × 104 cells로 24 시간 배양한 후, TY-NS-B를 농도별로 24시간 동안 처리하였다. 24시간 후 각각의 well에 1 ㎎/mL 농도의 MTT 용액을 50 μL씩 첨가하고 2시간 반응시킨 후, 상등액을 제거하고 DMSO를 100 μL씩 각각의 well에 첨가하여 녹인 후 UV/Visible spectrophotometer (Xma-3000PC, Human Corporation Co., Seoul, Korea)를 이용하여 540 ㎚에서 흡광도를 측정하였다.
SDS-PAGE 및 Western blot 분석
TY-NS-B가 처리된 세포에서 단백질을 추출하기 위해서 세포를 4℃에서 유지된 1 × phosphate-buffered saline (PBS)로 2회 세척한 후, protease inhibitor cocktail (Sigma-Aldrich Co.)과 phosphatase inhibitor cocktail (Sigma-Aldrich Co.)이 포함된 radioimmunoprecipitation assay buffer (Boston Bio Products, Ashland, MA, USA)를 4℃에서 30분간 처리하여 lysis시켜 단백질을 얻었다. 추출된 단백질은 Bicinchoninic acid protein assay (Pierce Biotechnology Inc., Waltham, MA, USA)로 정량분석 후, 동일량의 단백질을 SDS-acrylamide gel로 전기영동하고 PVDF membrane (Bio-Rad, Hercules, CA, USA)에 이동시킨 후 5% non-fat dry milk로 상온에서 1시간 동안 blocking하였다. 1시간 후, 1차 항체를 5% non-fat dry milk에 용해시켜 4℃에서 16시간 동안 반응시킨 후 membrane을 0.05% tween-20이 포함된 tris-buffered saline (TBS-T)로 5분간 3회 세척하였다. 그 후 2차 항체는 5% non-fat dry milk에 용해시켜 membrane에 상온에서 1시간 처리하였고, TBS-T로 5분간 3회 세척 후 membrane은 ECL western blotting substrate (Amersham Biosciences Co., Little Chalfont, England)를 이용하여 단백질을 확인하였다.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
TY-NS-B가 처리된 세포에서 total RNA를 추출하기 위해서 세포를 4℃에서 유지된 1 × PBS로 2회 세척한 후, RNeasy Mini kit (QIAGEN GmbH., Hilden, Germany)를 이용하여 total RNA를 추출하였다. 1 ㎍의 total RNA를 Verso cDNA synthesis kit (Thermo Fisher Scientific Inc., Waltham, MA, USA)를 이용하여 cDNA를 합성하였다. PCR은 PCR master mix kit (Promega Co., Madison, WI, USA)를 이용하여 수행되었고, 사용된 primer는 Table 1과 같다.
Table 1. Sequence of oligonucleotide primers used for RT-PCR
Transient transfection
Flag-tagged wild type β-catenin과 Flag-tagged S33Y β-catenin construct의 transient transfection은 PolyJet DNA transfection reagent (SignaGen Laboratories, Rockville, MD, USA)를 이용하여 수행되었다. 6-well plate에 배양된 A549 세포에 1 ㎍의 construct와 PolyJet DNA transfection reagent가 희석된 plasmid mixture를 처리하여 48시간 동안 배양시켜 실험에 사용하였다.
통계분석
모든 결과는 3 회 반복 측정 후 평균 ± 표준편차로 나타내었고, 처리간 유의성은 Student’s t-test로 검증하여 p-value 값이 0.05 미만일 때 통계적으로 유의하다고 판정하였다(Microsoft Exel 2010, Microsoft, Redmond, WA, USA).
결과 및 고찰
TY-NS-B의 A549세포에 대한 세포생육억제와 β-catenin 단백질 수준 감소 유도 활성
TY-NS-B의 비소세포성 폐암 세포인 A549의 세포생육 억제활성은 MTT assay를 이용하여 평가하였다. TY-NS-B는 A549세포의 생육을 유의적으로 12.5 ㎍/mL에서 9.4 ± 1.6%, 25 ㎍/mL에서 19.3 ± 2.5%, 그리고 50 ㎍/mL에서 58.7 ± 1.2%를 억제하였다(Fig. 1A).
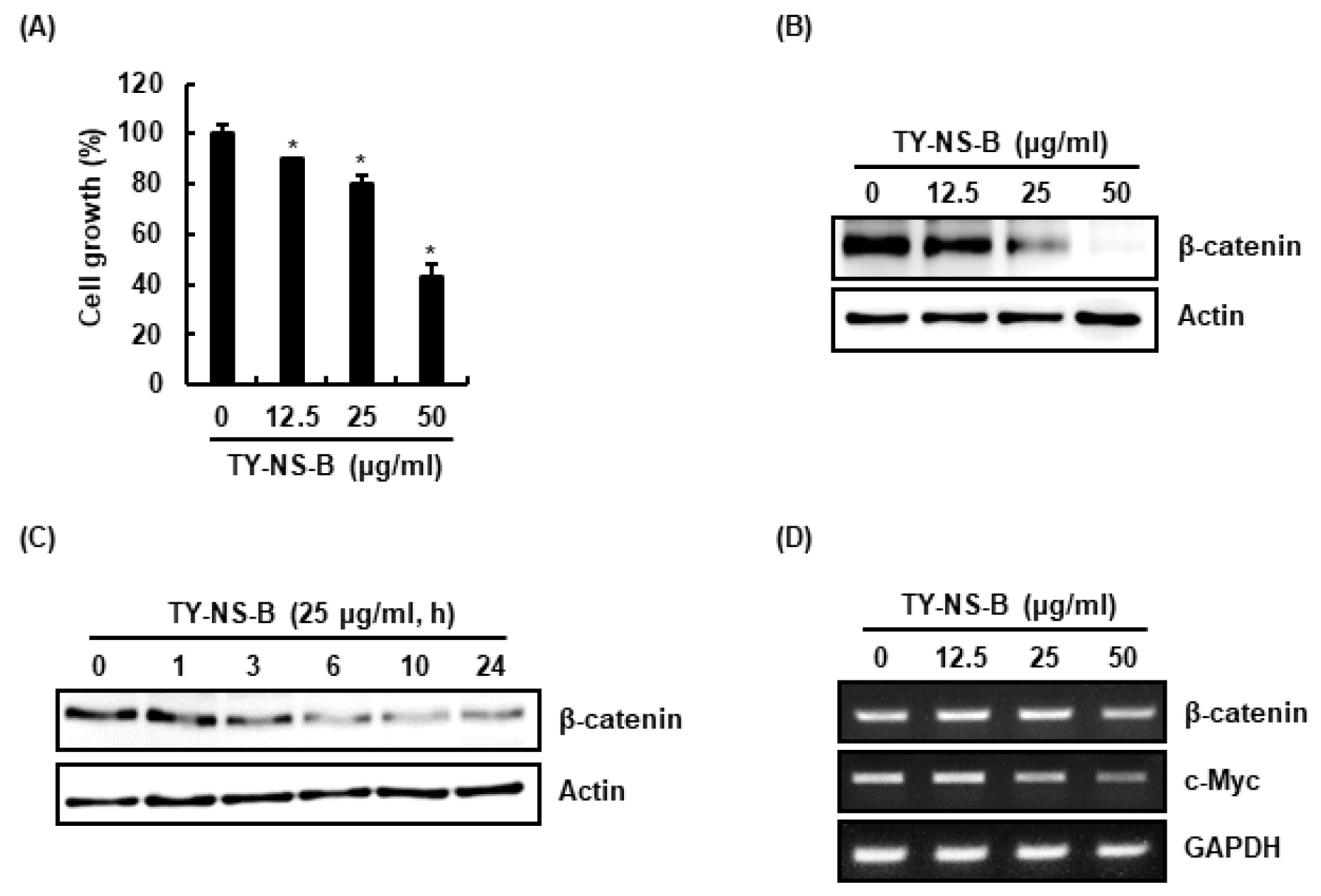
Fig. 1.
Effect of TY-NS-B on the cell growth and β-catenin expression in human lung cancer cells, A549. (A) Cell growth was evaluated by MTT assay at 24 h after TY-NS-B treatment. *p < 0.05 compared to cell without TY-NS-B. (B) A549 cells were treated with TY-NS-B at the indicated concentrations for 24 h. Western blot analysis was performed against β-catenin. Actin was used as internal control. (C) A549 cells were treated with TY-NS-B (25 ㎍/mL) for the indicated times. Western blot analysis was performed against β-catenin. Actin was used as internal control. (D) RT-PCR analysis of β-catenin and c-Myc gene expression was performed at 24 h after TY-NS-B treatment.
β-catenin은 암화과정의 악성형질 전환에 중추적 역할을 하는 Wnt 신호전달의 핵심 구성요소로 알려져 있다(Jin et al., 2017). 세포막, 세포질 및 핵에 존재하는 β-catenin은 세포 내 존재위치에 따라 이중적 역할을 한다(Li et al., 2014; Scanlon et al., 2013). 세포막에 존재하는 β-catenin은 cadherin과 결합하고 있어 암세포 간 부착과 암세포 성장을 억제하지만, 세포질과 핵에 존재하는 β-catenin은 Wnt 신호전달의 중심 분자로 작용한다(Li et al., 2014; Schmalhofer et al., 2009). Wnt 자극이 없을 때, Axin/APC/CK-1/GSK3β로 구성된 복합체는 β-catenin의 threonine-41(T41)/serine-33(S33)/serine-37(S37)을 인산화시켜 β-catenin의 분해를 유도하지만, Wnt 자극은 Axin/APC/CK-1/GSK3β로 구성된 복합체의 활성을 억제하여 β-catenin의 세포질 축적을 유도한다. 세포질에 축적된 β-catenin은 핵내로 전이되어 암세포의 생육과 관련된 c-Myc과 같은 유전자의 발현을 촉진시킨다(Tam and Weinberg, 2013). 그리하여 β-catenin은 항암제 개발을 위한 잠재적 표적단백질로 여겨지고 있다. 최근 β-catenin은 비소세포성 폐암 발생을 촉진시킨다고 보고되었다(Pacheco-Pinedo et al., 2011). TY-NS-B가 A549세포의 β-catenin 발현에 미치는 영향을 평가한 결과, TY-NS-B는 농도 의존적으로 β-catenin 단백질 수준을 감소시켰다(Fig. 1B). 또한 TY-NS-B는 처리 3시간부터 β-catenin 단백질 수준을 감소시켰다(Fig. 1C). TY-NS-B에 의한 β-catenin 단백질 수준 감소가 β-catenin의 전사활성 억제에 기인한 것인지 확인하기 위해, TY-NS-B 처리에 따른 β-catenin mRNA 변화를 측정하였다. 그 결과, TY-NS-B는 β-catenin mRNA의 발현에는 영향이 없었으나, β-catenin의 표적유전자인 c-Myc의 발현을 억제하였다(Fig. 1D). 본 결과를 미루어 볼 때, TY-NS-B는 β-catenin의 단백질 수준을 감소를 통해 세포생육과 관련된 c-Myc의 발현을 억제하여 A549세포의 생육을 억제하는 것으로 판단된다.
TY-NS-B의 GSK3β 비 의존성 β-catenin 단백질 분해 유도활성
TY-NS-B는 A549세포에서 β-catenin mRNA 발현에는 영향이 없었으나, β-catenin단백질 수준은 감소시키는 것을 확인하였다. 본 결과는 TY-NS-B에 의해 유도되는 β-catenin 단백질 수준 감소는 단백질 분해에 기인하는 것으로 가설된다. 사실 β-catenin 단백질 분해 유도를 통해 β-catenin 표적 발암유전자를 억제하여 항암활성을 나타내는 다양한 화합물들이 개발되고 있다(Hwang et al., 2016). 그리하여 TY-NS-B가 β-catenin 단백질 분해에 관여하는지 평가하기 위해 A549세포에 단백질분해 효소 억제제인 MG132를 2시간 전처리하고, TY-NS-B를 10시간동안 처리한 후 Western blot 분석을 통해 β-catenin 단백질 변화를 확인하였다. 그 결과, MG132가 처리되지 않은 A549세포에서는 TY-NS-B에 의해 β-catenin 단백질이 감소되었으나, MG132가 처리된 세포에서는 TY-NS-B에 의한 β-catenin 단백질 감소가 관찰되지 않았다(Fig. 2A). 본 결과로 미루어 볼 때, TY-NS-B에 유도되는 β-catenin 단백질의 수준감소는 β-catenin 단백질 분해에 기인하는 것으로 판단된다.
β-catenin 단백질의 분해는 GSK3β에 의한 S33, S37과 T41의 인산화에 의해 유도된다고 알려져 있다(Liu et al., 2002). 인산화된 β-catenin은 β-Trcp에 의해 인식되고 26S proteasome에 의해 빠르게 분해된다(Xu et al., 2010). 그리하여 TY-NS-B가 β-catenin 단백질의 인산화를 유도하는 확인한 결과, β-catenin 단백질의 인산화가 TY-NS-B 처리 1시간부터 증가하였다(Fig. 2B). 또한, β-catenin 단백질의 인산화를 유도하는 GSK3β가 TY-NS-B에 의한 β-catenin 단백질 분해에 관여하는지 확인한 결과, GSK3β가 처리된 세포와 처리되지 않은 세포에서 모두 TY-NS-B에 의해 β-catenin 단백질이 분해되는 것을 확인하였다(Fig. 2C). 본 결과를 미루어 볼 때, TY-NS-B에 의한 β-catenin 단백질 분해는 GSK3β 의존성 β-catenin 단백질 인산화에 기인하지 않는 것으로 가설된다. 이 가설을 검증하기 위해 Flag-tagged wild type β-catenin과 S33/S37T41이 돌연변이 된 Flag-tagged β-catenin을 형질전환 시키고, TY-NS-B를 처리 한 후, Western blot으로 Flag-β-catenin을 분석 한 결과, TY-NS-B에 의해 모두 Flag-β-catenin이 감소되는 것을 확인하였다(Fig. 2D). 따라서, 본 결과를 미루어 볼 때, TY-NS-B에 의한 β-catenin 단백질 분해는 GSK3β 의존성 β-catenin 단백질 인산화에 기인하지 않는 것으로 판단된다.
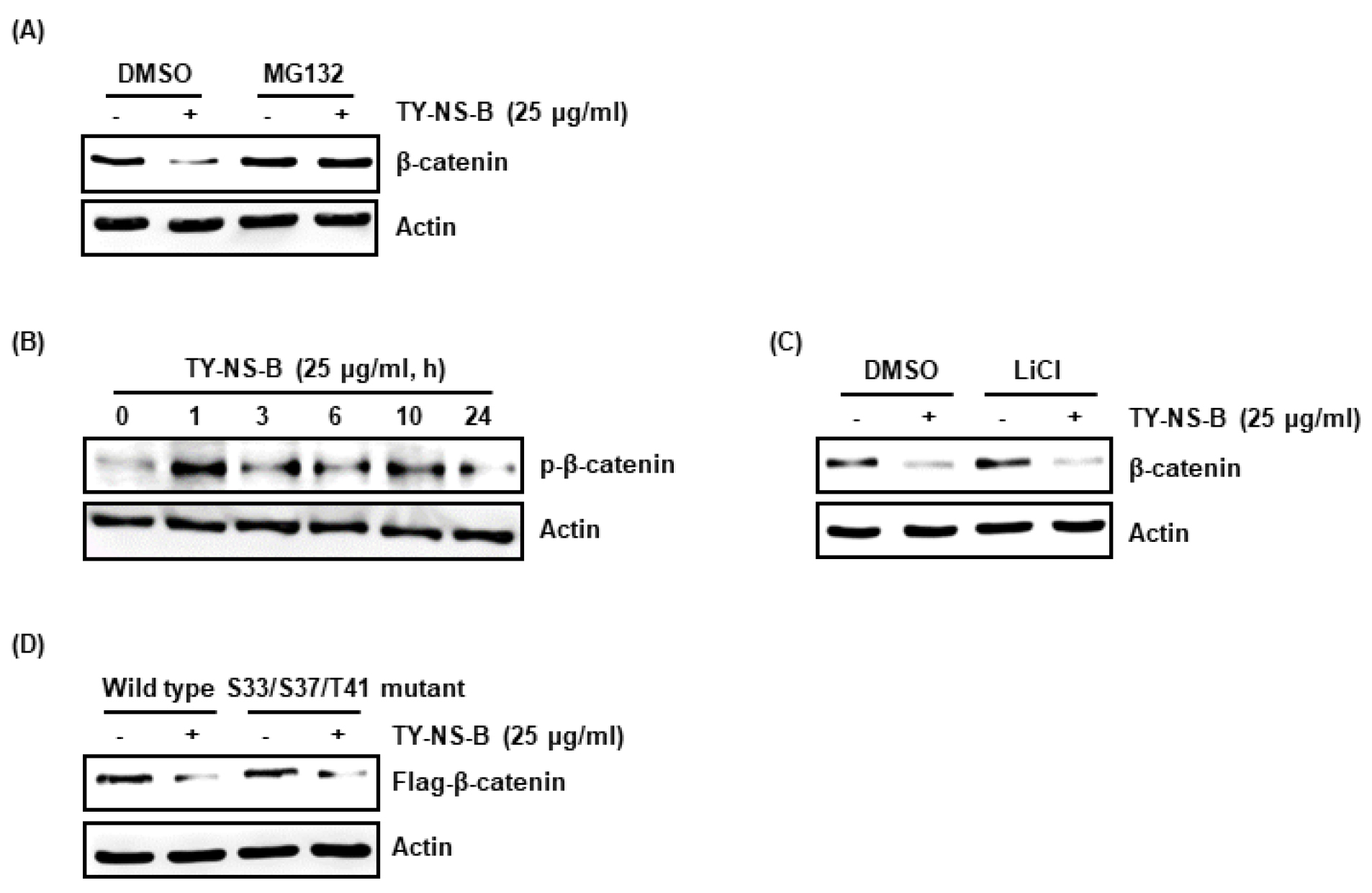
Fig. 2.
Effect of TY-NS-B on β-catenin proteasomal degradation in human lung cancer cells, A549. (A) A549 cells were pretreat with 20 μM of MG132 for 2 h and then co-treated with TY-NS-B for 10 h. (B) A549 cells were treated with TY-NS-B for the indicated times (C) A549 cells were pretreat with 20 mM of LiCl for 2 h and then co-treated with TY-NS-B for 10 h. (D) A549 cells were transfected with Flag-tagged wild type β-catenin or Flag-tagged S33/S37/T41 mutant β-catenin construct for 48 h and the co-treated with TY-NS-B for 12 h. Western blot analysis was performed against β-catenin. Actin was used as internal control.
적요
이상의 연구 결과로 미루어 볼 때, 참식나무 기주 참나무겨우살이(TY-NS-B)는 GSK3β에 의한 β-catenin 인산화에 의존하지 않는 β-catenin의 분해 유도를 통해 암세포의 생육과 관련된 c-Myc의 발현을 억제하며, 이것은 비소세포성 폐암의 생육억제와 관련이 있는 것으로 판단된다. 또한, 본 결과는 항암을 위한 대체보완소재로 참식나무 기주 참나무겨우살이의 활용이 가능할 것으로 판단된다. 그러나 추가적 연구를 통해 참식나무 기주 참나무겨우살이가 어떻게 β-catenin의 분해 유도에 관여하는지에 대한 기전연구와 관련 활성물질의 분석연구가 필요할 것으로 사료된다.
